|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Основные методы изучения эволюционного процесса ⇐ ПредыдущаяСтр 9 из 9
Рассмотрим главнейшие из методов изучения эволюционного процесса, представляемых биологическими дисциплинами в последовательности, которая отражает проникновение эволюционных идей в эти дисциплины: сначала палеонтологические, биогеографические, морфологические, эмбриологические и систематические, а затем данные генетики, биохимии, молекулярной биологии. Палеонтологические методы. По существу, все без исключения методы палеонтологии как науки об ископаемых организмах могут рассматриваться как методы изучения эволюционного процесса. Рассмотрим более подробно лишь главнейшие палеонтологические методы изучения эволюции: выявление ископаемых промежуточных форм, восстановление филогенетических рядов и обнаружение последовательности ископаемых форм. Ископаемые переходные формы – формы организмов, сочетающие признаки более древних и молодых групп. Поиски и детальные описания таких форм служат важными методами восстановления филогенеза отдельных групп. Яркий представитель переходных форм – ископаемая Ichthyostega (рис. 6.3), позволяющая связать рыб с наземными позвоночными. Наиболее древние наземные позвоночные из группы стегоцефалов также сохраняют некоторые рыбообразные черты.
Рис. 6.3. Примеры ископаемых переходных форм: А – переходная форма от рыб к наземным позвоночным – девонская ихтиостега (Ichthyostega), Б – прообраз переходной формы от рептилий к птицам – юрская первоптица (Archaeopteryx); В – переходная форма от рептилий к млекопитающим – звероподобная рептилия из группы терапсид (Lycaenops) (по И.И. Шмальгаузену. 1969; А. Ромеру, I968; P. Питерсону, 1978) Переходными формами от рептилий к птицам являются юрские первоптицы Archaeopteryx с длинным, как у рептилии, хвостом, несросшимися позвонками и брюшными ребрами, развитыми зубами. Но это были уже настоящие птицы: тело покрыто хорошо развитыми перьями, передние конечности превращены в типичные крылья. Последний из рассматриваемых в этой связи примеров – звероподобная рептилия Lycaenops из группы терапсид. Развитие большой зубной кости (os dentale), вторичного костного нёба, типичная для млекопитающих дифференцировка зубов на клыки, резцы и зарезцовые зубы, как и многие другие черты, делали общий облик этого животного похожим на хищных млекопитающих. Но по ряду основных черт строения и образу жизни это были настоящие рептилии. В настоящее время описаны и многие другие ископаемые переходные формы не только крупных (типы, отряды, классы), но и более мелких (отряды, семейства) групп животного и растительного мира. Исследование переходных форм – важный метод изучения эволюционного процесса. Палеонтологические ряды – ряды ископаемых форм, связанных друг с другом в процессе эволюции и отражающие ход филогенеза. По числу найденных промежуточных звеньев такой ряд может быть более или менее подробным, но во всех случаях это должен быть именно ряд форм, близких не только основными, но и частными деталями строения и, несомненно, генеалогически связанных друг с другом в процессе эволюции. Такие ряды ископаемых предков особенно интересно анализировать с эволюционной точки зрения в отношении современных форм: доступность для глубокого анализа заключительного, современного нам звена цепи позволяет лучше понять особенности развития всего эволюционного ряда. К настоящему времени известно немало палеонтологических рядов (например, лошадей, носорогов, слонов). Анализ эволюционного ряда лошадей (рис. 6.4) начат еще исследованиями основателя эволюционной палеонтологии В.О. Ковалевского (1842– 1883).
Рис. 6.4. Эволюционное древо семейства лошадиных (Equidae). Видны постепенная смена листоядных форм травоядными и сокращение числа пальцев от пяти до одного (по Дж. Симпсону, 1948, упрощенно): 1 – Eohippus; 2 – Miohippus; 3 – Parahippus; 4 – Pliohippus; 5 – Equus На примере анализа ствола лошадиных видна постепенность процесса эволюции: сменяющие друг друга ископаемые формы приобретали все большее сходство с современными. При сравнении эоценового эогиппуса с современной лошадью трудно убедительно доказать их филогенетическую связь. Наличие многих последовательно сменяющих друг друга форм на обширных пространствах ряда континентов (эволюция лошадей происходила в основном в Северной Америке и Евразии, хотя некоторые формы проникли и в Южную Америку) позволило построить филогенетический ряд с высокой степенью достоверности. На этом же примере видно, что, строго говоря, речь идет не просто о ряде форм, а о более сложных пространственно-временных отношениях групп. Развитие таких сменяющих друг друга пучков (вееров), форм чрезвычайно характерно для отдельных стволов любых групп в процессе эволюции. Последовательность ископаемых форм. Палеонтологические ряды конструируются на основании анализа отдельных разрозненных находок, относящихся к разным территориям. При этом всегда вероятно, что какие-то формы, жившие в прошлом, выпадают из-за отсутствия достаточно представительного материала. Однако современная палеонтология дает еще одно неопровержимое документальное свидетельство эволюционных процессов, протекавших в отдельных группах. При определенных благоприятных условиях сохраняются в ископаемом состоянии в одном и том же месте все вымершие формы. При послойном анализе таких отложений можно получить, истинную последовательность возникновения и изменения форм в эволюции (рис. 6.5).
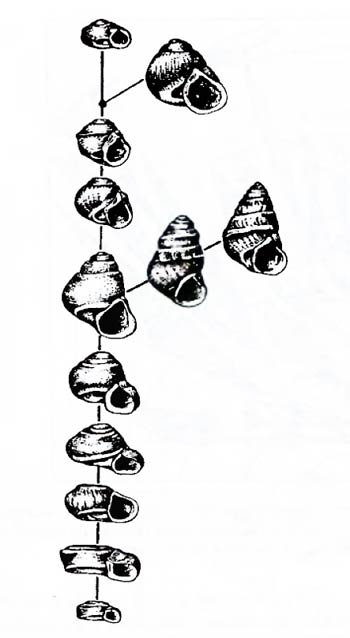
Рис. 6.5. Последовательность ископаемых форм верхнеплиоценовых моллюсков рода Gyraulus из Штайнхайма (Вюртемберг) (по И.И. Шмальгаузену, 1969) При анализе последовательности ископаемых форм возникает возможность определить реальную скорость протекания эволюционного процесса (подробнее см. гл. 15, 16). По поводу рядов ископаемых моллюсков И.И. Шмальгаузен (1884–1963) писал: «Вряд ли возможны еще более объективные доказательства существования эволюции».
Рис. 6.6. Увеличенный разрез ископаемого коралла. По числу суточных слоев роста в стенке скелетных пластинок было установлено, что в девоне год состоял из 400 дней, а к карбоне – из 390 (из М.Ф. Ивахненко и В.А. Корабельникова, 1987) Изучение смены флор и фаун, эволюции экосистем. В последние десятилетия в изучении ископаемых останков достигнуты большие успехи. Методы растворения окружающей породы позволяют восстанавливать даже очень тонкое строение вымерших микроскопических организмов (рис. 6.6). Применение палеомагнитных, палеохимических, радиоавтографических и других методов исследования позволяет выяснить особенности прошлых климатов и условий существования, например соленость того или иного участка океана, температуру воды, газовый состав атмосферы. Проведение более широких геологических и палеонтологических исследований дает исследователям все возрастающее число ископаемых форм (рис. 6.7). Все это существенно уменьшает «неполноту геологической летописи» (Ч. Дарвин), хотя, конечно, не ликвидирует ее полностью (табл. 6.1).
Рис. 6.7. Филогенетическое древо рептилий наглядно иллюстрирует значение изучения вымерших групп для восстановления филогенеза группы в целом. Современные крокодилы (и птицы!) оказываются потомками прежде мощно развитой группы археозавров (по данным разных авторов) Иногда возникает реальная возможность детальной реконструкции экосистем прошлого (рис. 6.8), выяснения их последовательной смены в процессе эволюции конкретных участков Земли. Такие последовательные реконструкции картин прошлой жизни являются мощными интегрировавшими методами изучения основных направлений развития жизни на Земле (см. также гл. 4, 19).
Рис. 6.8. Материал для реконструкции экосистем прошлого: основные микроископаемые верхнего докембрия Учуро-Майского района, Восточная Сибирь (по А.Ф. Вейсу, 1993) Являясь ведущими при изучении процессов макроэволюционных (текущих обычно в масштабах сотен тысяч – сотен миллионов лет), палеонтологические методы не могут быть применены, как правило, при изучении микроэволюционных процессов – во временных масштабах сотен и тысяч лет. Биогеографические методы. Биогеография дает в руки исследователей методы, позволяющие проанализировать общий ход эволюционного процесса в самых разных масштабах. Таблица 6.1. Оценка полноты современных палеонтологических данных по некоторым группам (по данным разных авторов из М. Ридли, 1985)
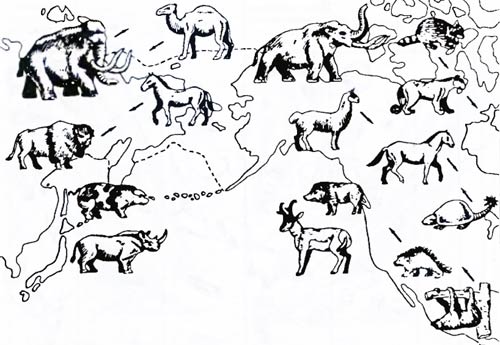
Сравнение флор и фаун. О масштабах эволюции, связанных с возникновением целых флор и фаун, можно говорить на основании изучения особенностей развития современных континентов Земли (см. рис. 6.1) в сопоставлении с анализом их населения. В конце триасового периода Пангея начала раскалываться на два сверхконтинента: северный – Лавразию, южный – Гондвану. Гондвана, в свою очередь, разделилась на Южноамерикано-африканский континент, Австралию вместе с Антарктидой и огромный остров, который много лет спустя «причалит» к Азии, образуя Индостан (см. рис. 6.1). В конце юрского периода Лавразия начинает раскалываться на Северную Америку и Евразию. Но Южная Америка и Африка (вместе с Мадагаскаром) еще образуют монолит. Следы этого былого единства сохраняются в современной фауне. Таковы ящерицы – игуаны Мадагаскара и Южной Америки, многочисленные сомовые и харациновидные рыбы Южной Америки и Африки. Ближайший родственник африканской шпорцевой лягушки (Xenopus) (строго говоря, это не настоящая лягушка) – суринамская жаба (Pipa), вынашивающая на спине развивающиеся икринки. А вот южноамериканских парадоксальных жаб, у которых, как у живущих у нас чесночниц, головастик больше взрослой особи, и эндемичных пустынных жаб Австралии связывает гораздо более отдаленное родство. Это понятно – Антарктида с Австралией уже отделились (но в Австралию уже успели проникнуть предки австралийских сумчатых, похожие на ныне живущих американских опоссумов). В конце мела – начале палеоген-неогена расположение материков начинает приближаться к современному. Возникает Атлантический океан, пока еще не очень широкий (через него, видимо, на каких-то плавучих островках из Африки в Южную Америку перебираются примитивные обезьяны и древесные дикобразы). Начинает намечаться разрыв между Австралией и Антарктидой. Мадагаскар отрывается от Африки и становится заповедником древних форм. Индостан движется на север, чтобы в конце концов, столкнувшись с Азией, закрыть древнее море Тетис и породить Гималаи. Но Лавразия еще не разделилась окончательно, и обмен видами между Старым и Новым Светом продолжается. Проанализируем несколько примеров сходства и различия населения отдельных территорий с эволюционной точки зрения. Фауна млекопитающих, характерная для Восточной Европы, сохраняется в основном такой же и за Уралом – в Северной Азии. Это объясняется тем, что на всей огромной территории Северной Евразии нет особых преград для расселения крупных и средних по величине млекопитающих. Фауна млекопитающих Северной Африки очень близка к таковой Северной Азии, и различие не превышает, как правило, родового ранга. Для фауны Северной Америки, так же как и для фауны Северной Евразии, характерны такие млекопитающие, как лоси, куницы, норки, росомахи, белые медведи, пищухи, летяги, бурундуки, сурки, суслики и многие другие близкие формы. Сходство фауны рассматриваемых территорий определяется тем обстоятельством, что сравнительно недавно существовал широкий «мост» между отделенными ныне континентами Евразии и Северной Америки – Берингия (рис. 6.9). Всего лишь миллион лет назад, как показывают палеогеографические данные, эти континенты были связаны друг с другом,
Рис. 6.9. Северная Америка и Азия неоднократно соединялись территорией Берингии, в связи с чем становился возможным широкий обмен фаун. Положение животных на рисунке соответствует направлению движения видов (по Дж. Симпсону, 1983) Сравним фауну млекопитающих Северной и Южной Америки. Несмотря на большую территориальную близость этих континентов, различия между фаунами оказываются огромными: если при сравнении фауны млекопитающих Северной Америки и Северной Евразии мы обнаруживаем в Северной Америке только три эндемичных семейства (табл. 6.2), то в Южной Америке – более 10, а число эндемичных родов достигает 80%. Только здесь живут ленивцы (Bradypodidae), муравьеды (Myrmecophagidae), броненосцы (Dasypodidae) – представители отряда неполнозубых млекопитающих. Не меньшие отличия и орнитофауны Южной Америки. Такое своеобразие животного населения этой части планеты объясняется тем, что на протяжении десятков миллионов лет Южная Америка была полностью изолирована от остальных континентов. Несколько раз возникавший узкий Панамский перешеек не мог обеспечить широкого обмена фауной. В сходном положении оказалась и Австралия: на протяжении более 120 млн лет она не соединялась с другими материками. За это время на австралийском материке самостоятельно, без влияния со стороны других фаун, развивались сумчатые и клоачные млекопитающие. В современной фауне Австралии можно выделить как бы четыре этапа вселения: на первом этапе туда проникли (или сохранились?) сумчатые (Marsupialia) и однопроходные (Monotremata), давшие к настоящему времени множество разнообразных видов, родов, семейств. На втором этапе, во время одного из сильных понижений уровня Мирового океана (несколько миллионов лет тому назад), по возникшему тогда сухопутному мосту из Южной Азии туда проникли некоторые грызуны. К настоящему времени эти грызуны представлены 26 видами мышей, а также шестью эндемичными родами (бобровые мыши, цепкохвостые крысы, тушканчикообразные крысы и др.). Третий этап вселения млекопитающих в Австралию – дикая собака динго (Canis dingo), попавшая на континент, видимо, вместе с человеком несколько десятков тысяч лет назад. Наконец, уже в наше время в Австралию с помощью человека попали и хорошо прижились кролики, олени и некоторые другие млекопитающие. Таблица 6.2. Характеристика основных фаунистических областей Земли
Выше мы остановились лишь на характеристике распределения высших позвоночных (в основном млекопитающих) по ряду зоогеографических областей Земли. Такой же анализ возможен и для растений (табл. 6.3). Все это показывает, как тесно связаны особенности видового состава отдельных районов планеты с историей этих территорий. Таблица 6.3. Характеристика основных флористических областей Земли
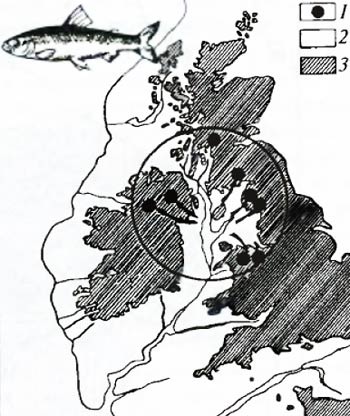
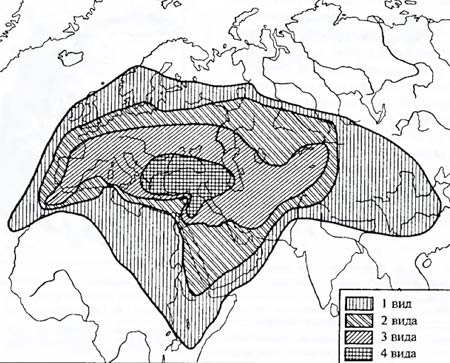
Рис. 6.10. Современное распространение пресноводных сигов рода Coregonus на Британских островах (по И.А. Бобринскому и др., 1946): 1 – распространение современных видов; 2 – бывшая территория суши; 3 – современная суша Особенности распространения близких форм. В некоторых реках Англии и Ирландии в настоящее время встречаются три вида пресноводных сигов рода Coregonus (рис. 6.10). Места их настоящего обитания показаны на карте кружочками. В недавнем прошлом вся Северо-Западная Европа была несколько приподнята над уровнем океана (по сравнению с современным положением). На месте Ирландского моря тогда должна была располагаться большая долина с огромным пресным водоемом, образованным стекающимися в эту долину реками. В этом водоеме и жил исходный прародительский вид современных сигов. Из-за проникновения морской воды в центральный водоем отдельные питающие его реки были изолированы друг от друга. На протяжении последующей самостоятельной эволюции группы изолированных сигов превратились в три разных вида. Этот пример показывает, как с помощью зоогеографических данных можно не только восстановить возможный ход эволюционного процесса, но и получить данные о темпах эволюции. Биогеографический анализ иногда позволяет выявить центры происхождения группы. На карте Евразии (рис. 6.11) нанесены ареалы всех видов коровяка Verbascum. По концентрации видов в Малой Азии можно сделать обоснованное предположение о существовании здесь центра не только разнообразия, но и происхождения всего рода.
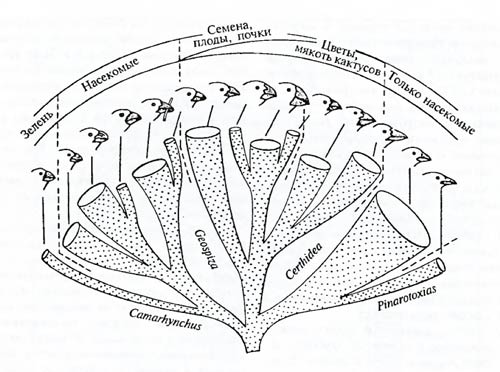
Рис.6.11. Ареал рода коровяков (Verbascum). Плотность штриховки пропорциональна числу видов, обитающих в данном регионе (из А.И. Толмачева, 1976) Островные формы. Фауна и флора островов оказывается тем более своеобразной, чем глубже и дольше эти острова были изолированы от основной суши. Так, например, сравнительно недавно потерявшие связь с материком Британские острова имеют фауну с небольшим числом автохтонных (возникших именно здесь) видов. К последним относятся куропатка-граус (Lagopus scoticus), два вида полевок, несколько уже упоминавшихся пресноводных сигов, ряд улиток и некоторое число видов мелких насекомых. С другой стороны, давно обособившийся (десятки миллионов лет) от африканского материка остров Мадагаскар имеет фауну настолько своеобразную, что выделяется в особую зоогеографическую подобласть: из 36 родов млекопитающих 32 рода эндемичны; из 127 родов птиц эндемична почти половина. Детальный анализ островной фауны позволяет восстановить пути эволюции группы близких видов. Со времени Ч. Дарвина классическим примером такого рода считается эволюция галапагосских вьюрков (Fringillidae). 13 видов этих вьюрков обитают ныне на Галапагосах и острове Кокос (в нескольких сотнях километров). Возраст Галапагосских островов – не более нескольких миллионов лет. Какой-то предок современных галапагосских вьюрков попал сюда, вероятно, задолго до других воробьиных птиц и обитал в изоляции от континентальных видов. Этот предковый вид освоил практически все адаптивные зоны (рис. 6.12). Один из предковых видов галапагосских вьюрков попал на остров Кокос, не образовав новых форм, поскольку, несмотря на разнообразие местообитаний, отдельные части видового населения не изолированы и могут широко скрещиваться.
Рис. 6.12. Дивергенция дарвиновых вьюрков (Geospizinae) на Галапагосских островах и о. Кокос (по Д. Лэку, 1949) Толщина стволов соответствует числу современных подвидов. Доля эндемиков растет по мере усиления изолированности островов (самое большое число эндемиков характерно для удаленных островов) В понятие «островных форм» должны быть включены и обитатели глубоких пещер, изолированных гор и водоемов. В пещерной фауне встречаются формы, появление которых хорошо объяснимо недавней эволюцией от родственных форм, живущих в окрестностях пещер. Так, например, ближайшие родичи обитающей в знаменитой Мамонтовой пещере в штате Кентукки (Северная Америка) слепой рыбы (Amblyopsis spelaea) – обычные обитатели водоемов этого района. С другой же стороны, пещерные обитатели могут иметь близкородственные формы чрезвычайно далеко (что косвенно указывает на исключительную древность вселения в пещеры). Так, например, ближайшие родственники европейского протея (Proteus anguinus) из подземных вод Югославии – амфибии из рода Necturus, обитающие в Северной Америке. Сходное положение складывается при обитании организмов в изолированных горных районах. Склоны Килиманджаро и Кении (Африка), разделенные всего тремястами километрами плоскогорья, имеют сходное население. 18 видам горно-лесных млекопитающих (обезьяны, белки, даманы и др.) со склонов горы Кения соответствуют 18 близкородственных, но все же иных видов со склонов Килиманджаро. Во всех случаях развития островных фаун и флор проявляется действие пространственной изоляции как эволюционного фактора (см. гл. 9). Много примеров анализа начальных стадий эволюции островных населений можно собрать и в наше время. При изменении биосферы человеком возникают новые водные бассейны, изолирующие ранее единые территории, появляются новые искусственные моря с островами разной величины и степени изоляции от окружающих материковых берегов. Все эти и подобные ситуации могут стать настоящей экспериментальной лабораторией в природе при изучении протекания начальных процессов эволюции. Сейчас возникло и интенсивно развивается новое направление биогеографии – островная биогеография (Р. МакАртур, Е.О. Вильсон), изучающая закономерности эволюции островных флор и фаун. Показано, что на каждом длительно существующем острове устанавливается равновесие в числе видов, попадающих туда из источников колонизации, и видов, вымирающих в результате небольших размеров острова и других факторов. Прерывистое распространение. Убедительными эволюционными «документами» служат факты так называемого прерывистого распространения организмов. В истории нашей планеты обычными были случаи, когда организмы, населявшие большие территории, исчезали на большей части прежнего ареала, не приспособившись к темпам изменения среды. Такие виды сохранялись лишь в некоторых ограниченных участках, имеющих условия, близкие к прежним. Примером такого рода для Европы является Альпийская фауна – комплекс видов, широко распространенных во время ледниковых периодов. Сейчас эти виды населяют лишь отдельные высокогорные участки Альп, Пиренеев и более низменные районы Северной Европы, Другой пример – майский ландыш (Convallaria majalis), широко распространенный по лесной зоне Европы. В лесах Кавказа обитает очень близкий вид C. transcaucasica, на Дальнем Востоке – C. Keiskei. Возможно, что прежде ландыш был широко распространен по всем лесным умеренным районам Северной Евразии, но во время наступления ледника сплошной ареал был разорван; ландыши сохранились лишь в немногих более теплых местообитаниях (Манчжурия, Закавказье, Средиземноморье). После отступления ледника отдельные части прежде единого вида стали развиваться самостоятельно. В Средиземноморье, на Кавказе и на территории Дальнего Востока во времена последних оледенений сохранялись теплолюбивые формы, вымершие на больших пространствах Евразии. Изучение таких биогеографических ситуаций позволяет познать особенности развития органического мира больших пространств и мелких участков. Реликты. О флоре и фауне далекого прошлого Земли свидетельствуют и реликтовые формы. Реликты – отдельные виды или небольшие группы видов с комплексом признаков, характерных для давно вымерших групп прошлых эпох. Среди животных одной из наиболее ярких реликтовых форм является гаттерия (Sphenodon punctatus) – единственный представитель целого подкласса рептилий. В ней отражены черты рептилий, живших на Земле десятки миллионов лет назад – в мезозое (рис. 6.13). Гаттерия живет в глубоких норах на островах залива Пленти (Новая Зеландия). Другой известный реликт – кистеперая рыба латимерия (Latimeria chalumnae), сохранившаяся малоизмененной с девона в глубоководных участках прибрежных вод Восточной Африки.
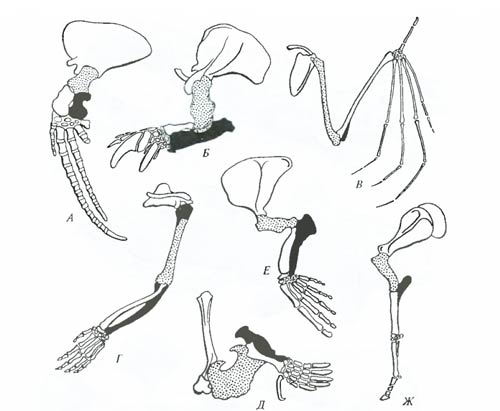
Рис. 6.13. Примеры реликтовых форм: А – гаттерия (Sphenodon punctatus); Б – латимерия (Latimeria chalumnae); В – опоссум (Didelphis marsupialis); Г – гинкго (Ginkgo biloba) (из И.И. Шмальгаузена, 1969; А. Ромера. 1969; Л.А. Курсанова и др., 1951) Среди растений реликтом может считаться гинкго (Ginkgo biloba), ныне распространенное в Китае и Японии только как декоративное растение. Облик этого растения дает нам представление о древесных формах, вымерших в юрском периоде. Эпоха существования Лавразии и Гондваны оставила много реликтовых форм с разорванным ареалом. Таковы осетровые, в первую очередь веслоносы (один вид в Китае, другой – в Миссисипи, этих пресноводных рыб теперь разделяют океаны), и целый ряд амфибий. Гигантская саламандра сейчас обитает в Китае (в Японию, по-видимому, завезена человеком), а ее ближайший родственник скрытожаберник – в Северной Америке. Но когда-то они населяли всю Лавразию: останки гигантской саламандры обнаружены в Швейцарии. Изучение реликтовых форм позволяет строить обоснованные предположения об облике давно исчезнувших групп, их образе жизни, условиях, существовавших миллионы лет назад. Таким образом, биогеографические методы дают возможность изучения как макроэволюционных процессов, так и процессов микроэволюции. В XX в. особенно возросло значение этих методов в изучении микроэволюции, что нашло свое выражение даже в возникновении специальных направлений динамической биогеографии (М. Удварди) и гено-(фено)географии (А.С. Серебровский). Морфологические методы. Использование морфологических (сравнительно-анатомических, гистологических и др.) методов изучения эволюции основано на принципе: глубокое внутреннее сходство организмов может показать родство сравниваемых форм. Гомология органов. Органы с общим планом строения, развивающиеся из сходных зачатков, находящиеся в сходном соотношении с другими органами и выполняющие как сходные, так и различные функции, называются гомологичными. Другими словами, это органы, имеющие глубокое сходство в строении, основанное на единстве происхождения соответствующих организмов. Различные по внешнему виду и функциям конечности млекопитающих состоят из сходных элементов: лопатки, костей плеча, предплечья, запястья, пясти, фаланг пальцев (рис. 6.14). Во всех случаях сохраняются единый план строения, сходства во взаимоотношении конечностей с остальными органами и онтогенетического развития.
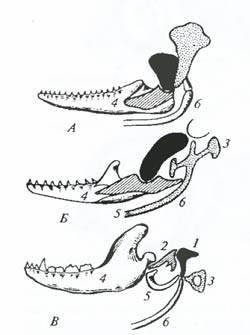
Рис. 6.14. Схема строения скелета передней конечности представителей разных отрядов млекопитающих: А – финвала (Balaenoptera physalus); Б – гигантского броненосца (Priudontes giganteus); В – рыжей вечерницы (Nyctalus noctula); Г – гориллы (Gorilla gorilla); Д – крота (Talpa europaea); Е – сивуча (Eumetopias jubatus), Ж – лошади Пржевальского (Equus przewalskii) (по И. Херань, 1968) Установление гомологии органов позволяет сделать вывод о родстве исследуемых организмов. Крупным успехом сравнительной анатомии в прошлом веке было, например, установление гомологии слуховых косточек позвоночных. Сравнение строения черепа в ряду низших и высших позвоночных (рис. 6.15) показывает, что у акуловых рыб центральная из костей – будущая наковальня – занимает в черепе важное конструктивное положение, являясь одной из мощных костей черепа. У костистых рыб она резко сокращается в размерах, сохраняя, однако, важное значение в конструкции черепа. У рептилий она резко изменена и служит рычагом при подвеске челюсти, а у млекопитающих занимает место в системе слуховых косточек.
Рис. 6.15. Гомология слуховых косточек среднего уха. Сравнительно-анатомический ряд позволяет восстановить эволюционный путь становления слуховых косточек млекопитающих: А – костистой рыбы; Б – пресмыкающегося; В – млекопитающего (из Н.Н. Иорданского, 1979): 1 – квадратная кость – наковальня млекопитающих; 2 – сочленовная кость – молоточек; 3 – гиомандибуляре – стремечко; 4 – зубная кость; 5 – угловая кость – барабанная кость млекопитающих; 6 – гиоид
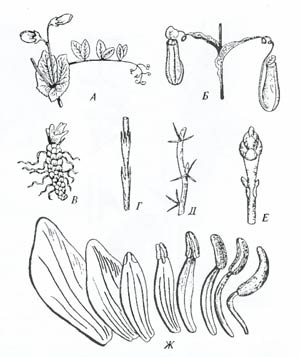
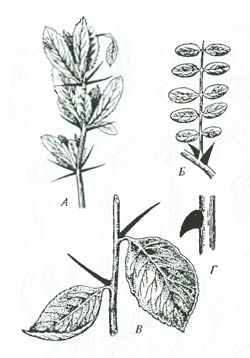
Рис. 6.16. Гомология некоторых органов у растений: А – перисто-сложный лист гороха с прилистниками и усиками; В – кувшинчики непентеса; В – чешуйки на корневище; Г – стеблевые чешуи хвоща; Д – колючки барбариса; Е – почечные чешуи. Все эти образования являются видоизменениями листовой пластинки. Постепенный переход от тычинок к лепесткам (Ж) в цветке белой кувшинки (Nymphaea alda) показывает путь происхождения тычинок (по И.М. Полякову, 1941; Ф.Н. Правдину, 1968) Пример гомологии у представителей царства растений приведен на рис. 6.16. Явление гомологии, или гомологического сходства, необходимо отличать от явления аналогии, или аналогичного сходства. Аналогичные органы лишь внешне сходны, что вызвано, как правило, выполнением сходных функций, а не общим происхождением. Для установления родства и выяснения путей эволюции изучаемых групп эти органы не имеют значения. На рис. 6.17 изображены колючки акации, барбариса, боярышника и ежевики. Все эти органы аналогичные и не свидетельствуют о родстве рассматриваемых форм, показывая лишь сходные направления приспособления, вызываемые в процессе эволюции действием естественного отбора (сохранялись и получали развитие формы растений, защищенные от поедания крупными растительноядными позвоночными).
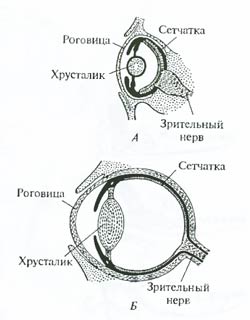
Рис. 6.17. Примеры аналогичных органов у растений: А – колючка барбариса обыкновенного (Berberis vulgaris), возникает из листьев; Б – колючка белой акации (Robinia pseudacacia)–из прилистников; В – колючка боярышника (Crataegus oxyacantha) – из побега; Г – колючка ежевики (Rubus caesius) – из коры (по Ф.Н. Правдину, 1968 с дополнениями) Иногда сходство аналогичных органов бывает столь глубоким, что для выявления их характера требуются специальные сравнительно-анатомические и эмбриологические исследования. Строение глаза наземных позвоночных и головоногих моллюсков, несмотря на удивительное сходство, является аналогичным (рис. 6.18). Глаз кальмара и глаз позвоночного развиваются из разных зачатков, сходство же определяется физической природой света. Недаром конструкция любого фотографического аппарата в точности повторяет строение глаза: хрусталик – объектив, радужина – диафрагма, сетчатка – чувствительная пленка, механизм аккомодации – наводка на резкость.
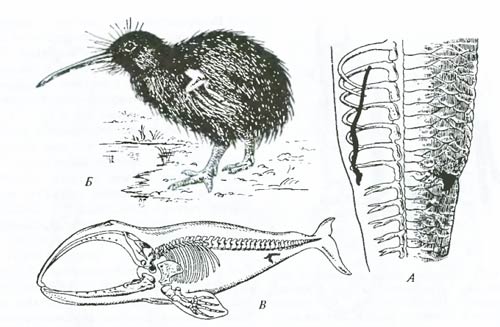
Рис. 6.18. Примеры аналогичных органов у животных. Сходство в строении глаза наземных позвоночных (А) и головоногих моллюсков (Б) основано на аналогии, поскольку органы развиваются из разных зачатков (из В. Даудесвелла, I960) Выявление характера сходства (гомология или аналогия) позволяет выяснить пути эволюционного развития, установить родство групп и, наконец, может показать направление действия естественного отбора. Рудиментарные органы и атавизмы. Данных гомологии недостаточно для восстановления пути развития той или другой группы, при этом трудно определить, например, какая из сравниваемых гомологичных форм родоначальная. В строении практически любого организма можно найти органы или структуры, сравнительно недоразвитые (лишенные каких-либо важных частей по сравнению с гомологичными структурами близких форм) и утратившие былое основное значение в процессе филогенеза; такие органы или структуры называются рудиментарными. Рассмотрим несколько классических примеров рудиментарных органов. У китообразных на месте заднего пояса конечностей располагаются в толще туловищной мускулатуры от одной до трех небольших косточек, связанных в настоящее время лишь с мьшцами мочеполовой системы (рис. 6.19). Эти рудименты тазовых костей подтверждают факт происхождения китов и дельфинов от наземных четвероногих предков с развитыми задними конечностями. Рудиментарные задние конечности питона также указывают на происхождение ныне безногих змей от предков с развитыми конечностями. У новозеландского нелетающего киви от крыльев остались лишь едва заметные выросты-рудименты, свидетельствующие, что у предков киви были настоящие крылья.
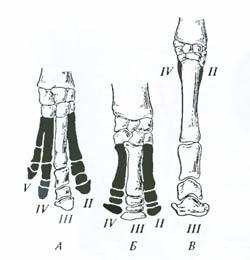
Рис. 6.19. Примеры рудиментарных органов: А – задние конечности питона (Python regius), Б – крылышко киви (Apteryx australis); В – элементы тазового пояса гладкого кита (Eubalaena glacialis) (по Ст. Сковрону, 1965; А.А. Парамонову, 1978) Много рудиментарных органов у человека. Это ушные мышцы и мелкая мускулатура, поднимающая основание волосяных фолликулов (у человека осталась лишь способность образования «гусиной кожи»). У диких млекопитающих поднимание волос и сейчас имеет важное терморегуляционное значение, у человека же эта функция кожной мускулатуры, несомненно, рудиментарна. Иногда рудиментарные органы могут достигать (у некоторых особей внутри вида) таких значительных размеров, что напоминают особенности строения предковых форм. Орган или структура, показывающие «возврат к предкам», называются атавистическими (от лат. attavus – предок). У человека атавизмами являются хвост, мощный волосяной покров на поверхности тела, наличие не двух, а нескольких пар сосков и т.п. Развитие у лошади вместо грифельных косточек (рудиментарных пальцев) настоящих боковых пальцев, так же как и развитие у китообразных выступающих на поверхность тела задних конечностей,– примеры атавизмов. Отличие рудиментарных органов от атавизмов состоит в том, что первые встречаются у всех членов данной популяции, вторые – лишь у немногих особей. Другую возможность разграничения рудиментов и атавизмов дает их разное функциональное значение. Каким бы незначительным ни был рудиментарный орган, он всегда выполняет в организме определенную функцию. Тазовые кости китообразных служат местом прикрепления мышц, обеспечивающих нормальную работу анального отверстия и половых органов, аппендикс у человека – органом лимфотворения, крыло у нелетающих птиц (например, страусов) – турнирным оружием и помогает птице при беге и т.п. Атавизмы же – все без исключения – не несут каких-либо специальных функций, важных для вида. Рудиментарные органы и атавизмы – убедительные доказательства процесса эволюции. Сравнительно-анатомические ряды. При рассмотрении гомологии органов речь шла о значении сравнения ряда специально подобранных организмов для изучения путей эволюции. Рассмотрим примеры эволюционного анализа сравнительно-анатомических рядов. На рис. 6.20 изображен ряд конечностей современных непарнокопытных млекопитающих: тапира, носорога, лошади, – показывающий путь эволюции, приведший к возникновению однопалой ноги у лошади. При переходе от обитания во влажных тропических лесах (тапир) к жизни в саванне (носорог) происходит редукция пальцев до трех. При переходе к жизни на открытых пространствах (лошадь) редукция пальцев идет еще дальше – до сохранения лишь единственного центрального пальца (с биомеханической точки зрения хождение на одном пальце чрезвычайно выгодно для обеспечения мгновенного отталкивания и быстрого бега). Сравнение современных однопроходных, сумчатых и плацентарных млекопитающих позволяет представить основной путь эволюции зверей – от откладывания яиц (ехидна и утконос) к рождению живых, но очень недоразвитых детенышей (сумчатые) и, наконец, к соединению организма зародыша с организмом матери (плацентарные).
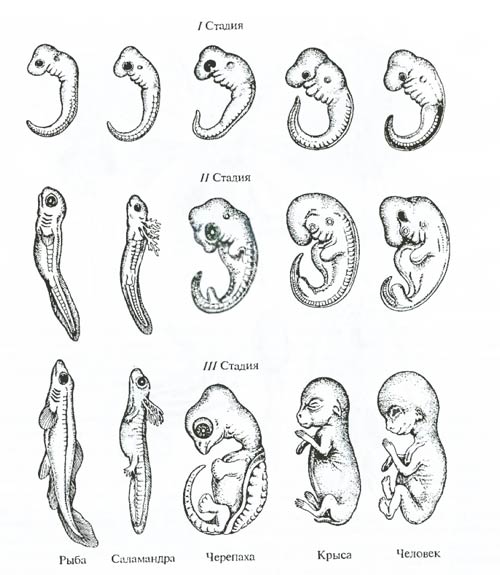
Рис. 6.20. Сравнительно-анатомический ряд: А – передние конечности тапира (Tapirus), Б – носорога (Ceratotherium); В – лошади (Equus); II–V– пальцы (из Д.К. Беляева и др., 1967) Морфологические методы изучения эволюции основаны на принципе «чем ближе родство, тем больше сходство». Они практически всегда сочетаются в эволюционном исследовании с другими подходами и методами (палеонтологическим, биогеографическим, генетическим и т.д.). Особенно тесное отношение к морфологическим подходам и методам изучения эволюции имеют данные эмбриологии. Эмбриологические методы. Эмбриология располагает целым арсеналом методов изучения эволюционного процесса. Среди них главные: выявление зародышевого сходства и изучение рекапитуляции. Выявление зародышевого сходства. В первой половине XIX в. выдающийся натуралист К. Бэр сформулировал «закон зародышевого сходства: чем более ранние стадии индивидуального развития исследуются, тем больше сходства обнаруживается между различными организмами. Например, на ранних стадиях развития эмбрионы позвоночных не отличаются друг от друга. Лишь на средних стадиях развития в сравниваемом ряду у зародышей появляются особенности, характерные для рыб и амфибий; на ее более поздних стадиях – особенности рептилий, птиц и млекопитающих (рис. 6.21).
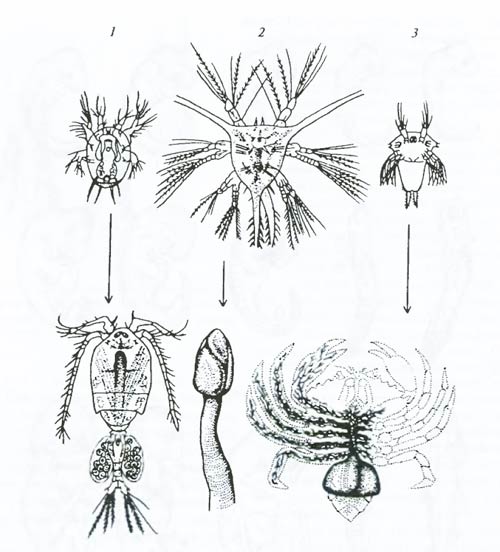
Рис 6.21. Явление зародышевого сходства. Эмбрионы всех позвоночных на ранних стадиях развития более сходны друг с другом, чем на более поздних стадиях (по Э. Геккелю, 1866). Размеры изображенных существ различны и некоторые детали опущены Вскоре после открытия К. Бэром Ч. Дарвин показал, что явление зародышевого сходства свидетельствует об общности происхождения и единстве начальных этапов эволюции сравниваемых форм. Принцип рекапитуляции. Явление зародышевого сходства позволило Ч. Дарвину и Э. Геккелю заключить, что в процессе онтогенеза как бы повторяются (рекапитулируют) многие черты строения предковых форм: на ранних стадиях развития повторяются признаки более отдаленных предков (менее родственных форм), а на поздних стадиях – близких предков (или более родственных современных форм). Все многоклеточные организмы проходят в развитии одноклеточную стадию, что указывает на происхождение многоклеточных от одноклеточных. Они проходят также стадию однослойного «шара» – ей соответствует строение некоторых современных простых организмов (например, вольвокса). Следующая стадия развития животных – обычно двухслойный мешок, или «шар»; этой стадии онтогенеза соответствует строение современных кишечнополостных (например, гидры). У всех позвоночных животных на определенной стадии развитии существует хорда (спинная струна); вероятно, у предков позвоночных хорда существовала всю жизнь. Строение личиночных форм нередко позволяет устанавливать родственные связи взрослых организмов, порой неузнаваемо различающихся. Для многих низших ракообразных характерна личинка науплиус с тремя парами конечностей и одним глазком (рис. 6.22). Взрослая форма непохожа на ракообразное, но науплиус отличается только роговидными выступами. Такие же «рожки» у науплиуса саккулины (Sacculina) (рис. 6.22, 3). Это усоногое во взрослом состоянии паразитирует в тканях высших ракообразных, прорастая в них корневидными выростами, полностью теряя облик ракообразного.
Рис. 6.22. Большое сходство науплиусов (планктонных личинок низших ракообразных) доказывает филогенетическое родство внешне непохожих взрослых форм (по В.H. Беклемишеву, 1964): 1 – веслоногий рачок (Calanus); 2 – усоногое ракообразное (Lepas); 3 – корнеголовый паразитический рачок (Sacculina) в теле краба Принцип рекапитуляции отнюдь не ограничивается лишь морфологическими изменениями. В процессе эволюции позвоночных происходит постепенная утрата ферментов, необходимых для распада мочевой кислоты (конечного продукта обмена пуринов). Так, у некоторых рептилий и птиц конечный продукт такого обмена – мочевая кислота, у земноводных и большинства рыб – мочевина, у беспозвоночных – аммиак. Эмбриобиохимические исследования показали, что зародыш птиц на ранних стадиях развития выделяет аммиак, на более поздних – мочевину, а на последних стадиях развития – мочевую кислоту. Сходным образом у головастиков – личинок бесхвостых амфибий – конечный продукт обмена – аммиак, а у взрослых лягушек – мочевина. Эти факты показывают на действие принципа рекапитуляции и среди физиолого-биохимических признаков. Концепция рекапитуляции помогает восстановить ход эволюционного развития многих групп и органов, палеонтологические материалы по которым отсутствуют или недостаточны. О современном состоянии этой концепции, о путях ее развития и уже известных ограничениях в применении этого метода исследования эволюции говорится в гл. 14. Методы систематики. Известно, что задача систематики – дисциплины, посвященной классификации организмов, – создание их естественной филогенетической системы. Поэтому выяснение систематического положения той или другой формы относительно других форм всегда связано с решением эволюционных проблем восстановления, генеалогии, путей эволюционного развития сравниваемых групп. Вклад теоретической систематики в эволюционное учение огромен. Перечислим лишь некоторые направления мысли: принципы стратиграфии, корреляции и иерархии признаков (Ж. Кювье), понятия родословного древа и филогенеза (Э. Геккель), адаптивная и инадаптивная специализация (В.О. Ковалевский), иерархия внутривидовых групп (подвид – раса – племя, А.П. Семенов-Тян-Шанский), концепция политипического вида (Э. Майр и др.), неравномерность темпов эволюции групп (Дж. Симпсон) и органов (О. Абель, Н.Н. Воронцов), уровни эволюции – грады и клады (Дж. Хаксли) и мн. др. Любое обстоятельное исследование систематики любой группы организмов в качестве основного результата имеет построение филогенетической системы – системы развития группы. Принцип рекапитуляции. Явление зародышевого сходства позволило Ч. Дарвину и Э. Геккелю заключить, что в процессе онтогенеза как бы повторяются (рекапитулируют) многие черты строения предковых форм: на ранних стадиях развития повторяются признаки более отдаленных предков (менее родственных форм), а на поздних стадиях – близких предков (или более родственных современных форм). Все многоклеточные организмы проходят в развитии одноклеточную стадию, что указывает на происхождение многоклеточных от одноклеточных. Они проходят также стадию однослойного «шара» – ей соответствует строение некоторых современных простых организмов (например, вольвокса). Следующая стадия развития животных – обычно двухслойный мешок, или «шар»; этой стадии онтогенеза соответствует строение современных кишечнополостных (например, гидры). У всех позвоночных животных на определенной стадии развитии существует хорда (спинная струна); вероятно, у предков позвоночных хорда существовала всю жизнь. Строение личиночных форм нередко позволяет устанавливать родственные связи взрослых организмов, порой неузнаваемо различающихся. Для многих низших ракообразных характерна личинка науплиус с тремя парами конечностей и одним глазком (рис. 6.22). Взрослая форма непохожа на ракообразное, но науплиус отличается только роговидными выступами. Такие же «рожки» у науплиуса саккулины (Sacculina) (рис. 6.22, 3). Это усоногое во взрослом состоянии паразитирует в тканях высших ракообразных, прорастая в них корневидными выростами, полностью теряя облик ракообразного.
Рис. 6.22. Большое сходство науплиусов (планктонных личинок низших ракообразных) доказывает филогенетическое родство внешне непохожих взрослых форм (по В.H. Беклемишеву, 1964): 1 – веслоногий рачок (Calanus); 2 – усоногое ракообразное (Lepas); 3 – корнеголовый паразитический рачок (Sacculina) в теле краба Принцип рекапитуляции отнюдь не ограничивается лишь морфологическими изменениями. В процессе эволюции позвоночных происходит постепенная утрата ферментов, необходимых для распада мочевой кислоты (конечного продукта обмена пуринов). Так, у некоторых рептилий и птиц конечный продукт такого обмена – мочевая кислота, у земноводных и большинства рыб – мочевина, у беспозвоночных – аммиак. Эмбриобиохимические исследования показали, что зародыш птиц на ранних стадиях развития выделяет аммиак, на более поздних – мочевину, а на последних стадиях развития – мочевую кислоту. Сходным образом у головастиков – личинок бесхвостых амфибий – конечный продукт обмена – аммиак, а у взрослых лягушек – мочевина. Эти факты показывают на действие принципа рекапитуляции и среди физиолого-биохимических признаков. Концепция рекапитуляции помогает восстановить ход эволюционного развития многих групп и органов, палеонтологические материалы по которым отсутствуют или недостаточны. О современном состоянии этой концепции, о путях ее развития и уже известных ограничениях в применении этого метода исследования эволюции говорится в гл. 14. Методы систематики. Известно, что задача систематики – дисциплины, посвященной классификации организмов, – создание их естественной филогенетической системы. Поэтому выяснение систематического положения той или другой формы относительно других форм всегда связано с решением эволюционных проблем восстановления, генеалогии, путей эволюционного развития сравниваемых групп. Вклад теоретической систематики в эволюционное учение огромен. Перечислим лишь некоторые направления мысли: принципы стратиграфии, корреляции и иерархии признаков (Ж. Кювье), понятия родословного древа и филогенеза (Э. Геккель), адаптивная и инадаптивная специализация (В.О. Ковалевский), иерархия внутривидовых групп (подвид – раса – племя, А.П. Семенов-Тян-Шанский), концепция политипического вида (Э. Майр и др.), неравномерность темпов эволюции групп (Дж. Симпсон) и органов (О. Абель, Н.Н. Воронцов), уровни эволюции – грады и клады (Дж. Хаксли) и мн. др. Любое обстоятельное исследование систематики любой группы организмов в качестве основного результата имеет построение филогенетической системы – системы развития группы. Переходные формы. Несмотря на то, что между крупными естественными группами животных, растений и микроорганизмов, как правило, существуют глубокие разрывы, вызванные вымиранием промежуточных форм, в ряде случаев мы обнаруживаем переходные формы. Существование форм, сочетающих в своем строении признаки разных типов организации и занимающих поэтому промежуточное систематическое положение, определяется общим родством организмов. При таком родстве между отдельными, далеко отошедшими друг от друга крупными ветвями древа жизни могут существовать мелкие ветви, носящие промежуточный характер (рис. 6.23).
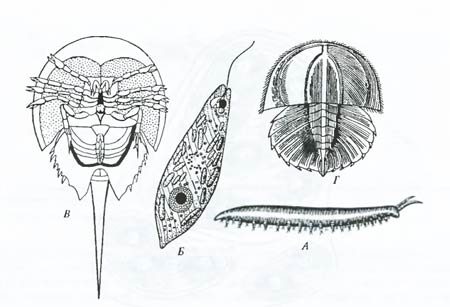
Рис. 6.23. Примеры ныне существующих форм: А – перипатус (Peripatus laurocerasus), несущий признаки членистоногих и кольчатых червей; Б – эвглена (Euglena viridis), соединяющая признаки животных и растений; В – мечехвост (Limulus polyphemus), занимающий промежуточное положение между современными типичными членистоногими и ископаемыми трилобитами; Г – личинка мечехвоста, похожая на личинку трилобита (по И.И. Шмальгаузену, 1969) Одним из примеров переходных форм между оболочниками (наиболее примитивной группой хордовых) и позвоночными животными служит род ланцетников, для которых характерны все основные признаки хордовых, но развиты они незначительно (Л.О. Ковалевский). Существование промежуточных форм в современном органическом мире – свидетельство единства организации крупных стволов древа жизни и единства их происхождения. Микросистематика. Начиная с классических работ П.П. Семенова-Тян-Шанского в случае хорошо изученных групп видов возникала возможность выявления их внутривидовой структуры – подвидов, рас и других группировок (микросистематика). Такое изучение имеет важное значение для понимания особенностей микроэволюции. Сейчас ясно, что в системе вида могут существовать сложные иерархические взаимоотношения популяций и их групп. При этом выявление истинного филогенетического родства таких группировок, необходимое для определения внутривидовых таксономических категорий, оказывается в то же самое время и восстановлением их микрофилогенеза – путей исторического развития отдельных частей видового населения и вида в целом (рис. 6.24).
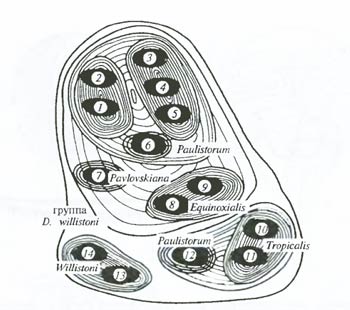
Рис. 6.24. Микрофилогенез дрозофил группы Willistoni можно восстановить на основании экспериментов по скрещиванию, анализа строения хромосом и биохимического полиморфизма. Иерархия филогенетических форм в данном случае должна быть описана так: популяция – подвид – полувид – вид – надвид (по Б. Спасскому и др., 1971) Популяционная морфология. А. Уоллес в XIX в. приводил вариационные ряды значений исследуемых признаков для небольших групп особей. С распространением популяционного мышления (см. гл. 7) морфологические исследования также оказались удобным оружием для изучения текущих процессов микроэволюции. Популяционно-морфологические методы, позволяют улавливать направления естественного отбора по изменению характера распределения значений признака в популяции на разных стадиях ее существования или при сравнении разных популяций (рис. 6.25, А, Б).
Рис. 6.25. Асимметрия в распределении значений количественного признака может показать направление давления естественного отбора (по А.В. Яблокову, 1966). Морфологические методы позволяют выделять среди однородной группы признаков те, которые находятся под большим или меньшим давлением со стороны отбора (рис. 6.26), хотя в этом случае само адаптивное значение признака может быть вскрыто лишь дополнительными эколого-физиологическими исследованиями. Одной из трудностей в применении морфологических методов к изучению процесса микроэволюции является сложность разграничения действия современных эволюционных факторов от изменчивости, определяемой широкой нормой реакций. Так, получивший одно время широкое распространение в нашей стране метод морфофизиологических индикаторов (С.С. Шварц), позволяющий различать физиологическое состояние популяций животных и направления ее адаптивной перестройки, часто оказывался скомпрометированным тем обстоятельством, что за эволюционно значимые индикаторы принимались мимолетные, кратковременные и обратимые изменения.
Рис. 6.26. Значения коэффициента вариации, резко выпадающие из «потока изменчивости» группы признаков, могут показывать на отношение данного признака к давлению отбора. В черепных промерах гренландского тюленя (Pagophilus groenlandica) пониженным значением коэффициента вариации отличаются размеры слуховой кости, а повышенной – носовой кости. Первый признак крайне важен и жестко контролируется отбором во все периоды онтогенеза, второй – малозначим (по А.В. Яблокову, 1966) Изучение микроэволюционного процесса с помощью морфологических методов является содержанием нового направления в современной морфологии – популяционной морфологии, дисциплины, тесно связанной с популяционной генетикой и экологией. Экологические методы. Экология, изучая условия существования и взаимоотношения между живыми организмами, играет важную роль в познании процессов эволюции. Весь эволюционный процесс является адаптациогенезом – процессом возникновения и развития адаптаций; экология вскрывает значение этих адаптаций. Например, экологи показали, что при колонизации островов виды с высокой скоростью размножения (r-стратегия, см. гл. 10) имеют больше преимуществ. На поздних стадиях колонизации по мере «насыщения» фауны и флоры преимущества получают виды с меньшими потенциями размножения, но обладающие высокой конкурентной способностью (K-стратегия). Эволюционные изменения хорошо прослеживаются и на примере взаимоприспособленности видов друг к другу, что играет важную роль в создании динамического равновесия и устойчивости экосистемы. В Центральной Америке и Мексике при отсутствии муравьев (Pseudomyrmex ferruginea), обычно поселяющихся колониями во вздутых шипах акации (Acacia cornigera), это дерево погибает из-за объедания ее листвы другими насекомыми. Бабочка-монарх (Danaus plexippus) делается несъедобной для хищников из-за накопления в теле при поедании листвы ядовитых растений высокотоксичных гликозидов. Примеров такого рода множество. Для обоснования теории естественного отбора чрезвычайно важными оказались опыты экологов на модельных популяциях по изучению роли окраски, поведения и формы тела у некоторых насекомых (см. гл. 11). Данные экологии позволяют уточнить и углубить доказательства эволюции из других областей биологии посредством выяснения роли конкретных адаптаций.
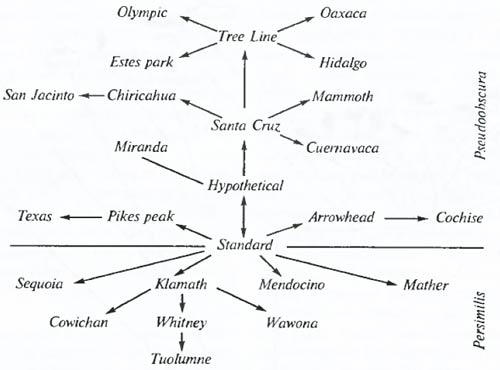
Рис. 6.27. Восстановление микрофилогенеза группы по инверсиям хромосом. Только в таком порядке могли возникнуть друг от друга обозначенные формы. Редкий случай надежного восстановления микрофилогенеза. Филогенетические взаимоотношения подвидов и видов в группе Drosophila pseudoobscura – D. persimilis, реконструириванные на основе распространения сложных инверсий в третьей хромосоме (по Ф.Г. Добржанскому. 1970) Генетические методы изучения эволюции разнообразны. Это и прямое определение генетической совместимости сравниваемых форм (например, посредством гибридизации), и анализ цитогенетических особенностей организмов. Изучением повторных инверсий в определенных хромосомах у разных популяций одного и того же или близких видов можно с большой точностью восстановить последовательность возникновения таких инверсий, т.е. восстановить микрофилогенез таких групп (рис. 6.27). Анализ числа и особенностей строения хромосом в группах близких видов часто позволяет выявлять направления возможной эволюции генома таких форм, т.е. выяснять их эволюционные взаимоотношения. На рис. 6.28 приведены хромосомные наборы ряда близких форм, которые могли возникнуть лишь в процессе последовательной полиплоидизации генома исходного типа, а в табл. 6.4– примеры рядов полиплоидных видов у некоторых растений.
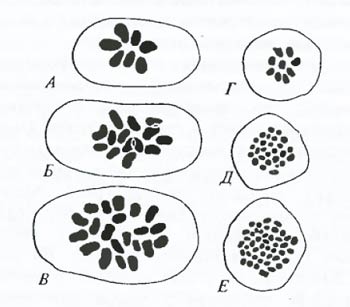
Рис. 6.28. Возникновение видов посредством полиплоидизации. А–В – хромосомные наборы в первой метафазе мейоза трех видов пшениц: Triticum aegilopoides, n = 7, Т. aestivum. n = – 21; T. dicoccoides, n = 14; Г–Е – то же, для видов хризантем: Chrysanthemum makinoi, n = 0; Ch. indicum, n = 18; Ch. japanense, n = 27 (по А. Мюнтцингу, 1967) Методологически важным результатом проникновения генетики в эволюционное учение стало широкое распространение экспериментальных подходов в решении эволюционных задач, а также применение точного (сравнительно с тем, что есть в других разделах биологии) аппарата генетических понятий. Таблица 6.4. Примеры полиплоидных рядов у некоторых покрытосеменных (по С.М. Гершензону, 1991)
Разработка современного эволюционного учения немыслима вне генетических подходов и методов. Развитие популяционной морфологии, фено- и геногеографии, микросистематики, кариосистематики и других «промежуточных» дисциплин (определяющих в известной мере современный этап развития эволюционного учения) наглядно иллюстрирует эту ведущую роль генетики. Методы молекулярной биологии. Уже Ч. Дарвин сделал успешные попытки использовать биохимические показатели для установления систематической принадлежности тех или иных форм. Однако только начиная с середины XX в. биохимические и молекулярно-биологические методы вышли, пожалуй, на передовые рубежи в изучении эволюционного процесса. По аналогии с изучением эволюции органов и онтогенеза в XIX – начале XX в. возникло целое направление изучения «молекулярной эволюции» (М. Кимура, В.А. Ратнер и др.). Здесь мы рассмотрим лишь некоторые из методов молекулярной биологии, широко применяющихся ныне для изучения эволюционного процесса. Выяснение строения нуклеиновых кислот и белков. На молекулярном уровне процесс эволюции связан с изменением состава нуклеотидов (в ДНК и РНК) и аминокислот (в белках). На современном этапе развития молекулярной биологии можно анализировать число различий в последовательностях элементов нуклеиновой кислоты или белка разных видов, судить по этому показателю о степени их отличий. Поскольку каждая замена аминокислоты в белке может быть связана с изменением одного, двух или трех нуклеотидов в молекуле ДНК, компьютерными методами можно вычислить максимальное и минимальное число нуклеотидных замен, необходимых для замещения аминокислот в белке.
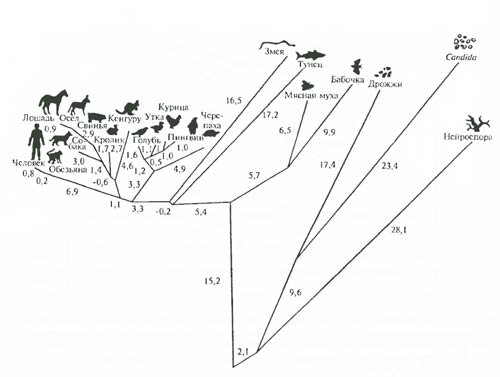
Рис. 6.29. Филогенетические дистанции между разными организмами, построенные на основании количественных изменений молекулярной структуры белка цитохрома C. Цифры – число мутаций (по А.Ф. Айяла, 1981) Получаемая таким образом информация поддается дальнейшей количественной оценке: при сравнении ряда организмов можно установить и степень различий (меру эволюционной дивергенции) макромолекул. Другое важное преимущество изучения эволюции методами молекулярной биологии – возможность сравнения сколь угодно далеких организмов – растений и животных, грибов и микроорганизмов. На рис. 6.29 показан результат изучения филогенетических отношений 20 разных организмов на основе определения минимального числа возможных нуклеотидных различий между генами, кодирующими синтез белка цитохрома C. Можно видеть, что в общем эти данные хорошо совпадают с выводами классической систематики. Сейчас построены многие сотни филогенетических деревьев макромолекул. При общем принципиальном совпадении с классическими эволюционными схемами эволюции крупных групп, молекулярными методами оказалось возможным вскрыть немало важных особенностей. Так, при анализе РНК, кодируемой не только ядерными генами, но и клеточными органеллами у растений, выяснилось, например (К. Вууз), что ядерная фракция p-РНК кукурузы (Zea mays) относится к ветви эукариот, а фракции из митохондрий и хлоропластов – к ветви эубактерий. Этот факт является мощным аргументом в пользу симбиотического происхождения эукариот: возникновения митохондрий от пурпурных бактерий, а хлоропластов – от цианобактерий (см. рис. 5.3). При реконструкции древа глобинов (В.А. Ратнер) удалось показать, что средняя скорость эволюции этих белков у животных заметно возрастала 400–500 млн. лет назад (период выхода позвоночных на сушу), когда глобин позвоночных приобрел тетрамерную структуру. Построение филогенетического древа генов, кодирующих синтез гемагглютининов H3 вируса гриппа показало, что скорость эволюции эпидемических вариантов этого вируса (испанка 1918–1919 гг., гонконгский грипп в середине – второй половине XX в. и др.) в несколько раз выше скорости эволюции неэпидемических штаммов. Аналогичные построения для вируса ВИЧ показали, что этот вирус очень близок к одному из вирусов обезьян. Он существовал в Центральной Африке до 1960 г., появился на о. Гаити в середине 70-х гг. и в США к 1978 г. Он распространяется ныне благодаря способности изменяться с невероятной скоростью – увеличивает агрессивность даже в теле одного человека на протяжении 1,5–2 лет. Молекулярный филогенетический анализ сегодня стал одним из главнейших методов выяснения филогенетических взаимоотношений самых разных групп живых существ и помогает решать немало спорных вопросов макроэволюции. Другим методом изучения эволюционного процесса на молекулярном уровне является оценка эволюционных изменений по степени сходства первичной структуры нуклеиновых кислот у различных групп организмов посредством гибридизации ДНК. Хотя около 90% ДНК не кодирует белков, тем не менее изучение ДНК позволяет оценивать филогению генов. Нуклеотидные последовательности позволяют судить об эволюции генов точнее, чем другие методы молекулярной биологии. Расщепленная на отдельные нити ДНК одного организма «гибридизируется» с молекулами ДНК другого вида и в зависимости от того, насколько различаются последовательности нуклеотидов, гибридизация захватывает большие или меньшие участки нитей ДНК. Этим достигается количественная оценка эволюционных изменений, происшедших со сравниваемыми видами. ДНК человека оказывается гомологичной ДНК макаки на 66%, быка – на 28, крысы – на 17, лосося – на 8, бактерии кишечной палочки – на 2%. Изучение особенностей эволюции на молекулярном уровне привело к идее молекулярных часов, не только отражающих, но и регулирующих эволюционный процесс. Такие расчеты основаны на предположениях о накоплении изменений в информационных макромолекулах с постоянной скоростью. Однако оказалось, что для растений, отличающихся в целом исключительно высокой скоростью эволюции первичной структуры ДНК, концепция молекулярных часов неприемлема: ДНК растений из разных семейств отличается в такой же степени, как ДНК животных разных классов. У растений в ходе эволюции очень быстро могут меняться и повторяющиеся, и уникальные последовательности нуклеотидов в молекулах ДНК. Скорость эволюции макромолекул в разных филогенетических линиях растений оказывается далеко не одинаковой. Так, темпы накопления изменений в нуклеотидных последовательностях у линий, ведущих к цветковым, более чем в 10 раз выше, чем в линии высших нецветковых. При изучении генов актинов было выявлено, что различия между близкими видами у растений превышают таковые разных классов у животных. Поэтому скорость хода молекулярных часов оказывается резко различной в разных группах, а также, по-видимому, в разные геологические эпохи (см. также гл. 15, 20). Иммунологические методы. Биохимия располагает специальными методами исследования, позволяющими с большой степенью точности выяснить «кровное родство» разных групп. При сравнении белков крови используется способность организмов вырабатывать (в ответ на введение в кровь чужих белков) антитела, которые затем можно выделить из сыворотки крови и определить, при каком разведении эта сыворотка будет реагировать с другой, сравниваемой сывороткой (реакции преципитации). Такой анализ показал, например, что ближайшие родственники человека – высшие человекообразные обезьяны, а наиболее далекие среди приматов – лемуры. Обнаружено серологическое сходство крови хищных млекопитающих с ластоногими, близость птиц к крокодилам и черепахам (птицы оказываются филогенетически ближе к крокодилам, чем крокодилы к змеям). Показаны глубокие серологические различия между хвостатыми и бесхвостыми амфибиями; найдены черты сходства по реакции преципитации между хордовыми и иглокожими и т.д. Подобные методы исследований применимы и к растениям. Биохимический полиморфизм. Начиная с 60-x годов в практику микроэволюционных исследований широко входит метод изучения вариаций белков, обнаруживаемых с помощью электрофореза (Р. Левонтин). С помощью этого метода можно с большей точностью, чем прежде, определять уровень генетической изменчивости в популяциях, а также степень сходства и различия между популяциями по сумме таких вариаций. Различия между популяциями по биохимическим показателям можно наглядно определять на карте (рис. 6.30) либо на дендрограмме, построенной по коэффициентам сходства и различий (рис. 6.31).
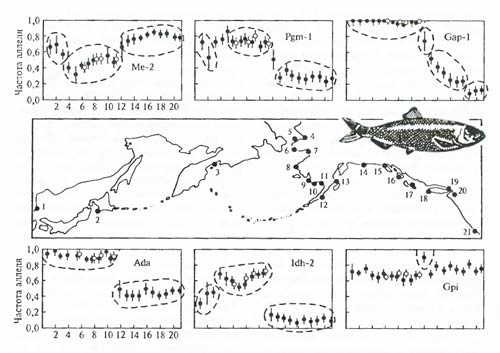
Рис. 6.30. Пример феногеографического анализа по частотам аллозимов (Ме-2, Pgm-l,Gap-l, Ada, Idh-2, Gpi) популяционной структуры тихоокеанской сельди (Clupea pallasi) (по В. Грант, Ф. Уттер, 1984). По частотам электроморф можно исследовать внутрипопуляционную пространственно-генетическую структуру и выявлять динамику этой структуры, т.е. исследовать начальные стадии процесса микроэволюции (см. гл. 8, 9).
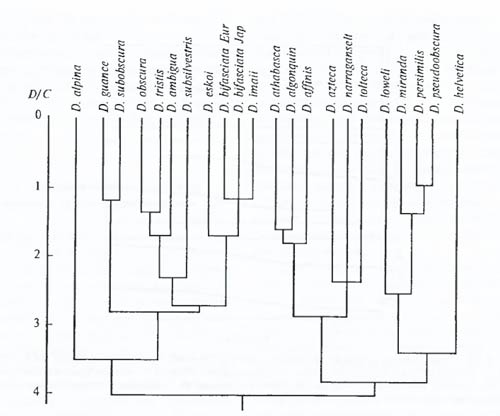
Рис. 6.31. Пример построения филогенетических отношений в виде дендрограммы на основании данных электрофореза белков: взаимоотношения между видами дрозофил группы D. pseudoobscura. D/C – среднее число аминокислот, по которым различаются ферменты разных видов (из С.М. Гершензона, 1979) При биохимических сравнениях ныне живущих организмов показано, что ранние метаболические звенья синтеза стеринов, каротиноидов, вторичных метаболитов, реакции гликолиза и дыхания у многих растений и животных сходны и не нуждаются в участии кислорода. Завершающие же стадии многих метаболических путей, особенно у высокоорганизованных существ, протекают с его участием. Это свидетельствует об их эволюционной молодости и сложении в аэробной фазе истории Земли. Таким образом, с помощью биохимических методов (и в том числе молекулярно-биологических) можно исследовать эволюционные процессы самого разного масштаба. Несомненно, что в ближайшие годы арсенал этих методов должен существенно расшириться, учитывая значительное развитие исследований в этой области. Методы моделирования эволюции. В последние десятилетия появилась возможность моделировать эволюционный процесс. Конечно, сложность организации жизни такова, что пока невозможно надежное моделирование развития экосистемы или целой группы, но влияние отдельных эволюционных факторов на популяцию и вид в целом поддается моделированию. Пока такое моделирование неизбежно связано с принятием целого ряда упрощающих условий (например, рассматриваются обычно лишь двулокусные генетические системы, принимается равновероятной возможность скрещивания особей в популяции друг с другом и другие упрощения, далекие от природных ситуаций). Но и при таких ограничениях моделирование иногда помогает уже сейчас решать ряд эволюционных задач. Моделирование эволюционного процесса позволяет, в частности, исследовать возможные влияния отдельных факторов на ход эволюции, их взаимодействие, выяснить в некоторых случаях возможные направления и результаты процесса эволюции (рис. 6.32). Математическое моделирование показало (А.Н. Колмогоров, 1935), что наибольшие темпы изменения популяций должны быть не в больших и не в малых изолированных популяциях, а в средних по величине, с периодически возникающими потоками генов между ними.
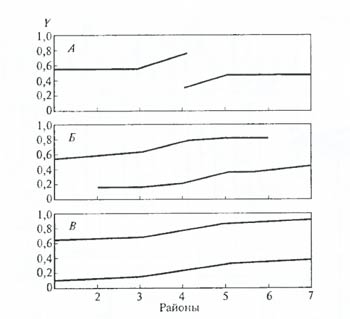
Рис. 6.32. Пример моделирования эволюции: распространение признака Y (форма рыла: острорылость – тупорылость) в популяциях ленка Brachymysrax в Азии: А – стадия расхождения признаков в районе первого контакта; Б – промежуточное состояние; В – возникшее стационарное состояние по данной паре признаков (по М.В. Мины, 1985) При применении быстродействующих ЭВМ с большой памятью возникает возможность как бы резко ускорить «процесс эволюции» и составить прогноз развития эволюционных событий при различном сочетании условий (см. также раздел 15.8). Особое внимание при моделировании процесса эволюции приходится уделять выбору необходимых и достаточных исходных предпосылок. Можно заложить в машину и «проиграть» различнейшие варианты взаимодействия тех или иных эволюционных сил в той или иной «обстановке», но если упустить какой-либо один из ведущих компонентов, то результаты моделирования могут получиться обратные реально существующим в природе. Определить набор необходимых и достаточных предпосылок для создания модели – дело очень сложное. Такие работы целесообразно организовывать только в тесном контакте математиков и биологов. Другие методы изучения эволюции. На основании данных, полученных сравнительной физиологией, можно проследить эволюционное развитие тех или иных функций. Так, по обмену веществ грибы ближе стоят к животным, чем к растениям (А.Н. Белозерский), что служит еще одним основанием для выделения их в самостоятельное царство. Значительный интерес для выяснения тонких особенностей протекания процесса эволюции представляет изучение поведения животных (этология), на что много внимания обращал Ч. Дарвин. У близких видов, недавно дивергировавших от общего предка, сохраняется (или рекапитулирует в процессе онтогенеза) множество общих мелких черт поведения (К. Лоренц, Н. Тинберген и др.). Некоторые характерные древние черты поведения у птиц и млекопитающих могут быть не менее важными показателями происхождения от общих предков, чем гомологичные органы или другие классические сравнительно-анатомические доказательства. В некоторых случаях эффективным оказывается использование паразитологического метода изучения эволюции. Многочисленными исследованиями доказано, что эволюция паразитов и хозяев протекает сопряженно. Известно также, что в некоторых группах паразиты оказываются специфическими для видов, родов или семейств. Поэтому по присутствию определенных паразитов порой можно с большой точностью судить о филогенетических связях видов – хозяев этих паразитов. По-видимому, любая биологическая дисциплина обладает своим специфическим набором методов изучения эволюционного процесса, и достаточно полное их перечисление практически невозможно в рамках одного курса. Эволюционная биология Эволюцио́нная биоло́гия — раздел биологии, изучающий происхождение видов от общих предков, наследственность и изменчивость их признаков, размножение и разнообразие форм в ходе эволюционного развития. Развитие отдельных видов обычно рассматривается в контексте глобальных преобразований флор и фаун, как компонентов биосферы. Эволюционная биология начала оформляться в качестве раздела биологии с широким признанием идей об изменчивости видов во второй половине XIX века. Эволюционная биология — междисциплинарная область исследований, поскольку она включает в себя как полевые, так и лабораторные направления различных наук. Вклад в эволюционную биологию вносят исследования в таких узкоспециальных областях, как териология, орнитология или герпетология, которые обобщаются для получения ясной картины развития всего органического мира. Палеонтологи и геологи анализируют окаменелости, чтобы получить сведения о темпах и формах эволюции, а популяционная генетика исследует эти же вопросы теоретически. Экспериментаторы используют селекцию дрозофил для лучшего понимания многих проблем эволюционной биологии, например эволюции старения. В 1990-х годах биология развития вернулась в эволюционную биологию после длительного забвения в виде новой синтетической дисциплины — эволюционной биологии развития. Эволюционная биология как академическая дисциплина стала общепризнанной в результате синтеза дарвиновской теории и генетики в 1930-х и 1940-х годах XX века. Основу новой теории заложили работы Четверикова[1], Фишера[2], Райта[3] и Холдейна[4], в которых рассматривалось воздействие естественного отбора на частоту аллелей в популяциях. Характер этих работ был скорее теоретическим, чем экспериментальнообоснованным[5]. Ситуацию исправила монография Феодосия Добжанского «Генетика и происхождение видов»[6]. В основу проблемы автор положил экспериментальную популяционную генетику. Теоретическая работа предшествующих авторов сопоставлялась с данными по изменчивости и отбору, полученными в ходе различных экспериментов. Добжанский полагал, что макроэволюционные процессы могут быть объяснены в терминах микроэволюции, которые протекают достаточно быстро, чтобы человек мог наблюдать их в экспериментах или в природе. Генетические идеи проникли в систематику, палеонтологию, эмбриологию, биогеографию. Из названия книги Джулиана Хаксли «Evolution: The Modern synthesis»[7] в научную литературу проник термин «современный синтез», обозначивший новый подход к эволюционным процессам. Выражение «синтетическая теория эволюции» в точном приложении к данной теории впервые было использовано Джорджем Симпсоном в 1949 году. Эта теория стала основой для развития эволюционной биологии во второй половине XX века. Подавляющее число новых идей в этой области рождалось из дискуссий вокруг синтетической теории, причём как из её защиты, так и из критики. |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 1401; Нарушение авторского права страницы