|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Ретротранспозоны группы Gypsy
В последние годы активно изучались ДКП-ретротранспозоны группы Gypsy. Эти элементы имеют одну либо две открытые рамки считывания. Из них самым изученным является элемент gypsy, его структура изображена на рисунке 1.3.
Рис. 1.3 Структура ретротранспозона gypsy Благодаря наличию функционально активной третьей ОРС gypsy , в отличие от других МГЭ, обладает инфекционной способностью. Продуктом третьей ОРС gypsy, как и у ретровирусов, является белок, участвующий в процессе узнавания клеточных рецепторов и проникновения вируса в клетку (Song et al., 1994). Было доказано, что gypsy может передаваться в результате микроинъекций плазмы эмбрионов из линии с активной транспозицией gypsy в линии без транспозиций. Было установлено, что gypsy может инфицировать мух через прямой контакт или через пищу (Kim et al., 1994a). Таким образом, было показано, что gypsy способен к горизонтальному переносу. Ретротранспозон gypsy был отнесен к, так называемым «эррантивирусам»- эндогенным ретровирусам беспозвоночных. Это первый ретровирус, обнаруженный у беспозвоночных животных. Транспозиция gypsy может вызывать мутации в генах ovo , cut , su (Hw) и др.(Prud ’ homme , 1995). Ген ovo , расположенный в Х хромосоме и необходимый для развития яичников у Drosophila , является горячей точкой инсерции ретротранспозонов группы gypsy и copia (Mevel-Ninio et al ., 1989). Эти мутации являются точечными и приводят к формированию нефункционального продукта. (Mevel-Ninio et al ., 1991). Для данной мутации характерно явление реверсии. В частности реверсия может происходить в результате транспозиции МГЭ gypsy . Ген cut так же является горячей точкой транспозиции gypsy. Инсерция в этот ген ведет к формированию слабых обрезанных крыльев у мухи. Другие ретротранспозоны группы Gypsy представлены на филогенетическом древе, изображенном на рисунке 1.4.
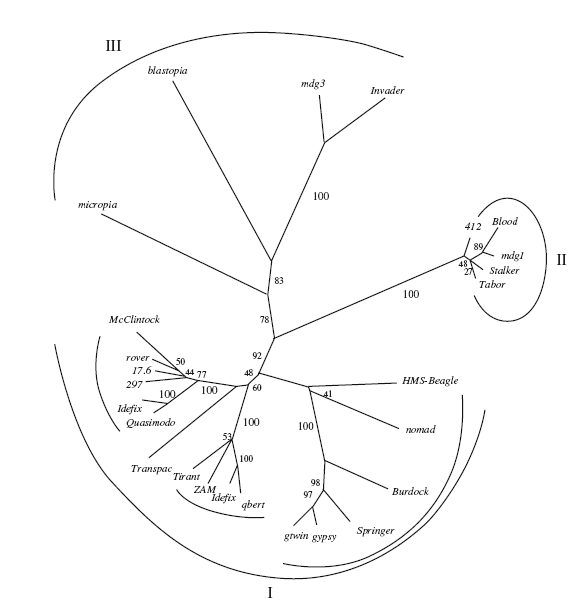
Рис.1.4. Филогенетическое дерево ДКП-ретротранспозонов группы gypsy D. m elanogaster . I. Элементы с двумя и тремя ОРС: McClintock , qbert , HMSBeagle, Burdock, Transpac , gypsy, ZAM, Idefix, Tirant, Quasimodo, nomad (opus), 17.6, 297, rover (pregun), springer, gtwin. II. Элементы с двумя ОРС: 412, Blood , mdg 1, Stalker , Tabor . III. Элементы с одной OPC: micropia , blastopia , mdg 3, Invader (Nefedova, Kim, 2009).
1.4.5. Генетический контроль транспозиции мобильных элементов группы gypsy . Высокая транспозиционная активность МГЭ опасна для организма, т.к. повышает частоту возникновения мутаций. По этой причине, эволюционно в геноме существуют механизмы, подавляющие транспозиции МГЭ. В линиях дикого типа частота транспозиций gypsy снижена, однако известны лабораторные линии, в которых частота транспозиции повышена. Значит, существует генетический контроль транспозиции МГЭ. Была получена линия MS D . melanogaster, в которой частота транспозиций gypsy увеличена, спонтанный мутагенез повышен до уровня 10-3-10-4 (Ким и др., 1989). Было сделано предположение, что генетический контроль gypsy в этой линии нарушен. Позже было показано, что контроль транспозиции gypsy осуществляется геном, flamenco, локализованном в 20А1-3 (Prud’homme et al ., 1995). Таким образом, линия MS – мутант по гену flamenco . Ген flamenco расположен на участке 20А1-3 между комплементационными группами extra organ ( eo ) и wings apart ( wap ) в β-гетерохроматине Х-хромосомы (Prud’homme et al ., 1995). В локусе flamenco присутствуют дефектные копии ретротранспозонов gypsy, Idefix и Zam (Pelisson et al. 1994; Robert et al. 2001; Desset et al. 2003). Регуляция контроля перемещения gypsy не затрагивает его промотор. Это было показано путем введения gypsy под другим промотором, сильно отличавшимся от исходного. Конструкция все равно подвергалась репрессии со стороны локуса flamenco . Позже было установлено, что подавление транспозиций МГЭ происходит по механизму РНК-интерференции Brenecke Malone et al. 2009 Brennecke, J., A.A. Aravin, A. Stark, M. Dus, M. Kellis, R. Sachidanandam, and G.J. Hannon. 2007. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128: 1089-103.] Malone, C.D., J. Brennecke, M. Dus, A. Stark, W.R. McCombie, R. Sachidanandam, and G.J. Hannon. 2009. Specialized piRNA Pathways Act in Germline and Somatic Tissues of the Drosophila Ovary. Cell.
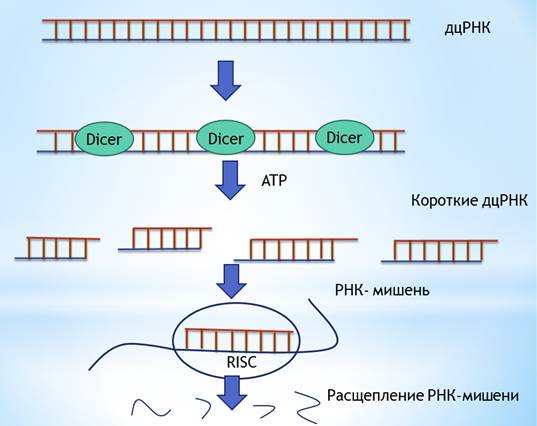
(ССЫЛКА). РНК-интерференция – механизм подавления экспрессии генов на посттранскрипционном уровне, вызванный появлением в клетке гомологичной двухцепочечной РНК (дцРНК). Процессы РНК-интерференции обнаружены в клетках многих эукариот, они играет важную роль в защите клеток от вирусов, МГЭ. Схема «общей» РНК-интерференции представлена на рисунке 1.5.
Рис. 1.5. Общая схема РНК- интерференции. В большинстве случаев, запуском механизма РНК-интерференции является дцРНК, которая может быть введена в клетку извне, образоваться в результате двунаправленной транскрипции гена, а в случае некоторых вирусов, появиться на определенных этапах их жизненного цикла. Далее дц-РНК процессируется РНКой III Dicer, и короткие РНК включаются в состав РНК-белкового комплекса, который узнает РНК – мишень и осуществляет ее деградацию. Этот комплекс называется RISC, от английского –RNA-induced silencing complex. Известны случаи, когда антисмысловым РНК предшественником является одноцепочечная РНК. Именно это было продемонстрировано в случае гена flamenco. (ссылка) Длинный антисмысловой транскрипт, считываемый с этого гена, процессирутся белком Piwi (cccc). Показано, что мутация в в гене piwi приводит к аккумуляции транскриптов Gypsy и других ретротранспозонов, что в результате может способствовать повышенной транспозиционной активности МГЭ. Таким образом, МГЭ группы Gypsy играют существенную роль в функционировании генома D . melanogaster. В результате своей транспозиционной активности, особенно на фоне мутаций в генах, ответственных за ограничение данного процесса, они могут нарушать работу многих клеточных генов, а значит, являться потенциальными факторами, влияющими на продолжительность жизни. II .МАТЕРИАЛЫ И МЕТОДЫ Материалы. В эксперименте использовались плодовые мухи Drosophila melanogaster (Meigen, 1830). Линии мух, используемые в эксперименте, и их характеристики представлены в таблице 2.1. Табл. 2.1 Линии мух и их характеристики.
Методы |
Последнее изменение этой страницы: 2019-04-21; Просмотров: 275; Нарушение авторского права страницы