|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Половое созревание и физиологическая зрелостьСтр 1 из 68Следующая ⇒
Половое созревание – это этап в жизни индивида, когда налаживаются эндокринные взаимоотношения в системе гипоталамус – гипофиз – яичники и устанавливается половая циклика, что сопровождается резким увеличением поступления в кровь половых стероидов. Возникает эндокринный фон, что, в свою очередь, обеспечивает интенсивный рост половых органов и молочной железы, ускоренное увеличение массы тела животного, формирование вторичных половых признаков; происходит развитие таза и других экстерьерных черт, присущим зрелым особям, формируется материнская доминанта. Началом полового созревания считают возраст, при котором у самки впервые проявились течка и охота как результат циклической деятельности яичников; это в свою очередь, означает приобретение способности к оплодотворению и плодоношению. С практической точки зрения важно знать, в каком возрасте молодые самки вступают в фазу полового созревания. Это, в частности, позволяет во-время изолировать их от самцов, чтобы избежать преждевременной беременности. С другой стороны, дает ориентир для выбора возраста и срока осеменения. У телок средний возраст вступления в фазу полового созревания составляет 9 мес. Однако диапазон различий бывает весьма значительным – от 8 до 18 мес. Это обусловлено рядом факторов: направление продуктивности, порода, условия выращивания, климат, перенесенные в раннем возрасте болезни. Относительно ранний возраст полового созревания отмечается у специализированных мясных пород (герефордская, абердин-ангусская и др.), поздний – у молочных пород (голштино-фризская, англерская, айрширская); комбинированные породы (симментальская, швицкая и др.) по данному признаку занимают промежуточное положение. Промышленное скрещивание ускоряет наступление этой фазы; очевидно, сказывается эффект гетерозиса. Интенсивное выращивание племенных телок, ориентированное на среднесуточный прирост живой массы 600-850 г, существенно ускоряет начало функционирования половой системы. Давно замечено, что выращенный в жарком климате молодняк отличается большей скороспелостью. У телок, перенесших в раннем возрасте ту или иную болезнь, в дальнейшем наблюдается отставание в росте и развитии, соответственно запаздывают сроки полового созревания и наступления физиологической зрелости. Не случайно зарубежные фермеры предпочитают не использовать таких животных для ремонта собственного стада, а передают в специализированные фермерские хозяйства на доращивание и откорм. Ремонтные свинки вступают в фазу полового созревания в возрасте 6-7 мес, овцы и козы в 7-10 мес, лошади в 15-24 мес. В фазу полового созревания происходит аутотренинг половой системы; ее неподготовленность проявляется нерегулярными половыми циклами, овуляцией без признаков течки и охоты и другими отклонениями. Окончательная стабилизация наступает в третий – четвертый половой цикл. Негативные последствия преждевременного наступления беременности могут выражаться в трудных или патологических родах, пониженной жизнеспособности приплода, малоплодии (свиньи), возникновении длительного или постоянного бесплодия из-за травм родовых путей, послеродовых осложнений. Под физиологической зрелостью следует понимать достижение самкой готовности к вступлению в репродуктивный период. Физиологическая зрелость наступает в следующие сроки: крупный рогатый скот – 12-18 мес, овцы – 12-13, свиньи – 10-12, лошади – 30-36 мес. На сроки наступления физиологической зрелости большое влияние оказывают направление продуктивности, порода, условия выращивания, индивидуальные особенности. Рекомендуемые отечественной зоотехнической наукой схемы выращивания ремонтных телок рассчитаны на достижение ими физиологической зрелости к 18-24-месячному возрасту, что предопределяет экстенсивный путь развития отрасли. Зарубежные технологии, напротив, ориентированы на интенсивное выращивание ремонтного молодняка. Стандартом для молочных и комбинированных пород в настоящее время принято считать возраст осеменения 15 месяцев; возраст первого отела – 24 месяца, при живой массе (для голштинов) 545 кг. Такая практика оправданна экономически, так как позволяет снизить стоимость выращивания на 20-35% по сравнению с отелом в 30-36 мес; оплата корма на 21-34% выше. Помимо этого, снижается потребность в первотелках, более полно реализуется генетический потенциал их продуктивности. Последнее объясняется тем, что беременность наслаивается на фазу интенсивного роста животного. В этом случае повышенная продукция гормона роста (соматотропина) усиливает действие плацентарных стероидов на формообразовательные процессы в молочной железе. Использование современных технологий кормления и содержания позволяет осеменять телок специализированных мясных пород в 13-14-месячном возрасте, при соблюдении условий: масса тела не менее 340-350 кг, упитанность заводская (чуть выше средней). Интенсивная технология свиноводства, в частности, предусматривает осеменение ремонтных свинок в 10-11-месячном возрасте, при массе тела 115-120 кг. Таким образом, определяющим критерием наступления физиологической зрелости, а следовательно пригодности молодой самки для вступления в репродуктивный период является живая масса – она должна составлять 65-70% таковой взрослых животных данной породы. Фолликулогенез Термин означает процесс развития фолликулов, охватывающий промежуток времени от момента выхода из латентного состояния первичного (примордиального) фолликула до овуляции. Закладка первичных фолликулов происходит еще во внутриутробный период. Начало потенциальным яйцеклеткам — оогониям дает зачатковый эпителий, который покрывает яичник снаружи. Непосредственно под зачатковым эпителием располагается тонкая белочная оболочка. Зачатковый эпителий — это морфогенно активная клеточная структура. Согласно современным представлениям, клетки зачаткового эпителия происходят из эктодермы желточного мешка. В эмбриональный период клетки эктодермы мигрируют и после колонизации поверхности яичника начинают быстро размножаться. В результате пролиферации и дифференциации зачаткового эпителия образуются оогонии. Последние мигрируют через белочную оболочку яичника в поверхностный слой корковой зоны. За счет пролиферации клеток мезенхимы вокруг оогония образуется однорядный слой эпителиоидных (фолликулярных) клеток. Новое образование носит название примордиальный (первичный) фолликул.
Рис. 6. Закладка примордиальных фолликулов: а, б – образование оогоний; в- миграция оогоний в корковую зону яичника; г – формирование первич ного фолликула
Первичные фолликулы располагаются под белочной оболочкой небольшими группами (гнездами). У телочки закладка первичных фолликулов наиболее интенсивно происходит на 110-130-е сутки внутриутробного развития. К моменту рождения общее их число достигает 75000. Принято считать, что гаметогенез (образование половых клеток) происходит лишь во внутриутробный период. Данную точку зрения авторы аргументируют тем, что с возрастом число первичных фолликулов непрерывно уменьшается; у старых коров их насчитывается всего лишь около 2500. Согласно другой точке зрения (Г.А. Черемисинов и др.), первичные фолликулы образуются на протяжении всей жизни самки; возрастное уменьшение их числа авторы объясняют атрофией и фиброзом соединительнотканых элементов коркового (генеративного) слоя яичника. С наступлением половой зрелости часть первичных фолликулов выходит из латентного состояния и вступает в фазу роста, при этом они преобразуются вначале во вторичные, затем в третичные фолликулы. Превращение первичного фолликула во вторичный начинается митозом эпителиальных (фолликулярных) клеток. В результате число клеточных слоев фолликулярного эпителия постепенно увеличивается и к концу фазы достигает 18-20. Одновременно фолликулярные клетки из плоских преобразуются вначале в кубические, затем в высокоцилиндрические; последние начинают секретировать фолликулярную жидкость. Островки жидкости постепенно сливаются и формируют полость фолликула. Расположенные вокруг растущего фолликула соединительнотканые клетки коркового слоя яичника находятся в морфогенно активном состоянии, обеспечивая формирование наружного слоя — капсулы. Таким образом, вторичному фолликулу присущи такие признаки: фолликулярная зона, состоящая из многорядного секреторного эпителия (гранулеза); капсула (тека) в фазе формирования; щелевидная полость, в которую вдается яйценосный бугорок (кумулюс) с расположенным в центре формирующимся яйцом (рис.7 ).
Вопреки традиционным представлениям, превращение первичного фолликула во вторичный — длительный процесс; так, у крупного рогатого скота и овец его продолжительность составляет около 130 суток. Полностью сформировавшийся вторичный фолликул вступает в фазу третичного фолликула, которая длится 34-43 суток. В течение этого срока завершается формирование капсулы и происходит ее дифференциация на наружный и внутренний слои. Полость становится доминирующей частью фолликула; она заполнена фолликулярной жидкостью, богатой эстрогенами и другими биологически активными веществами (простагландин Ф2α , коллагеназа, фертилизин и др.). Яйценосный бугорок отделяется от гранулезы, но сохраняет с ней связь посредством клеточного стебля (рис.8).
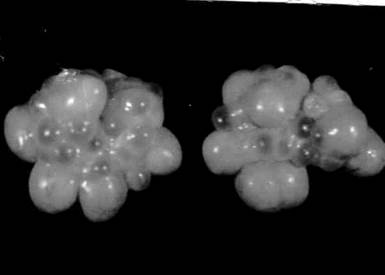
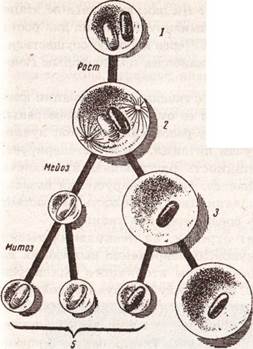
Рис. 8. Яичник половозрелой свиньи (фото) Третичный фолликул по мере роста приближается к поверхности яичника; он представляет собой тонкостенный пузырек диаметром 1-3 мм. Достигнув поверхности яичника, третичный фолликул начинает быстро расти за счет накопления фолликулярной жидкости и становится преовуляторным фолликулом (граафов пузырек). Это конечная (терминальная) фаза фолликулогенеза, длительность которой составляет 3-4 суток. К концу этой фазы фолликул достигает 16-18 мм в диаметре и возвышается над поверхностью яичника в виде купола. В центральной части купола стенка фолликула истончается, что в дальнейшем облегчает ее разрыв. Оогенез Одновременно с фолликулогенезом происходит оогенез. Этим термином обозначают превращение оогония в зрелое яйцо. Оогенез протекает в несколько фаз. В процессе превращения первичного фолликула во вторичный оогоний растет, вокруг него формируется из мукополисахаридов прозрачная оболочка. Фолликулярные клетки, прилегающие к прозрачной оболочке, принимают упорядоченное (радиальное) расположение и формируют клеточную оболочку потенциального яйца — лучистый венец. В цитоплазме клетки идет накопление питательных веществ в виде желточных шаров. В результате перечисленных изменений к концу первой фазы фолликулогенеза оогоний превращается в ооцит I порядка. По завершении фазы роста ооцит I порядка претерпевает мейотическое разделение, при этом один набор хромосом абстрагируется за пределы ядра в виде полярного тела, а вновь образованная клетка, имеющая гаплоидный набор хромосом, называется ооцитом II порядка.
Рис. 9. Схема оогенеза: 1 — оогоний, 2 — ооцит I порядка, 3 - ооцит II порядка, 4 — зрелое яйцо, 5 — полярные тела
После разрыва фоликула в яйцепровод поступает не зрелое яйцо, как принято считать, а ооцит II порядка. Заключительная фаза овогенеза имеет место уже в процессе оплодотворения, причем ее инициирует проникший в цитоплазму клетки спермий. Она состоит в том, что ооцит II порядка претерпевает второе мейотическое деление, с образованием зрелого яйца и полярного тела.
Атрезия фолликулов Под атрезией понимают деградацию растущего фолликула. Атрезии могут подвергаться все категории фолликулов. Посредством атрезии происходит саморегуляция их численности. На заключительном этапе фолликулогенеза она оптимизирует условия для роста доминантного фолликула. Через атрезию осуществляется ранний отбор яиц с наиболее полноценным генетическим материалом. Атрезия начинается с гибели и десквамации клеток гранулезы, отслоения ее от базальной мембраны. Вслед за этим происходит рассеивание клеток лучистого венца; не получая питания, ооцит дегенерирует. Фолликулярная жидкость рассасывается, полость фолликула уменьшается. После разрушения базальной мембраны в фолликулярную полость прорастает вновь образованная соединительная ткань. Как результат атрезии, из нескольких тысяч первичных фолликулов, одновременно вышедших из латентного состояния, фазы вторичного фолликула достигают 150-250, а к началу преовуляторной фазы в яичнике коровы их остается всего лишь 3-5. Финальная фаза отбора проявляется в том, что овулирует, как правило, один фолликул, остальные претерпевают атрезию. На протяжении жизни коровы овулирует около 50 фолликулов, что составляет лишь 0, 002% от общего их запаса в яичниках.
Овуляция Разрыв зрелого фолликула с выделением в брюшную полость ооцита II порядка вместе с окружающими его клетками лучистого венца и фолликулярной жидкостью называется овуляцией. Овуляции предшествуют лизис стенки фолликула под воздействием содержащегося в фолликулярной жидкости фермента коллагеназы и разрыв базальной мембраны с последующим проникновением кровеносных сосудов из внутреннего слоя капсулы в гранулезу (визуально отмечается гиперемия дна фолликула). Окончательное созревание фолликула и разрыв его стенки находятся под непосредственным контролем гормона передней доли гипофиза — лютропина. Так, овуляция происходит через 24-36 ч после поступления в кровь лютропина. У сельскохозяйственных животных овуляция спонтанная. Исключение составляют крольчихи, у которых овуляция рефлекторная, т. е. она наступает под воздействием полового акта.
Образование и функция жёлтых тел После того как произошла овуляция, полость фолликула заполняется кровью. Кровь свертывается, обеспечивая тампонаду дефекта и предотвращая дальнейшее ее поступление из поврежденных сосудов дна фолликула. За счет пролиферации и трансформации клеточных элементов стенки фолликула формируется в желтое тело которое постепенно замещает кровяной сгусток. При этом из наружного слоя капсулы внутрь врастают соединительнотканые тяжи (трабекулы);. пространство между ними заполняется секреторными клетками — лютеоцитами.
Рис. 10. Ооцит II порядка после выхода из полости фолликула
Различают два типа лютеоцитов: малые, берущие начало из клеток внутреннего слоя капсулы фолликула, и большие, являющиеся продуктом трансформации клеток гранулезы, причем преобладает вторая популяция клеток. В цитоплазме лютеоцитов накапливаются красящие пигменты каротин и ксантофилл, вследствие чего желтое тело приобретает яркооранжевый цвет. Замещение кровяного сгустка структурами желтого тела завершается на 5-е сутки после того как произошла овуляция. Дальнейшие структурные изменения характеризуются увеличением размеров и васкуляризацией желтого тела. У коров и телок пик развития и функциональной активности желтого тела приходится на 9-16-е сутки. Если беременность не наступила, то на 17-е сутки после овуляции желтое тело вступает в фазу регрессии. При этом оно быстро уменьшается в размере, становится плотным, изменяет цвет, Однако полное исчезновение морфологических структур желтого тела завершается лишь к концу следующего полового цикла. Таким образом, желтое тело полового цикла проходит три фазы: пролиферации, васкуляризации, регрессии (инволюции). По физиологическому назначению это временная железа внутренней секреции, основной ее продукт — стероидный гормон прогестерон. Половой цикл У самок сельскохозяйственных животных половая функция имеет циклический характер. На протяжении полового цикла в репродуктивных органах и организме самки протекает ряд взаимосвязанных процессов, направленных на создание благоприятных условий для транспорта половых клеток —спермиев и яиц, оплодотворения и развития беременности. Половой цикл принято рассматривать как стадийный процесс. Еще в 1900 г. английский биолог Хип предложил дифференцировать половой цикл на 4 стадии: проэструс, эструс, метэструс, анэструс. Проэструс — это период подготовки половых путей и организма самки к плодотворному осеменению. Эструс (от греческого слова «неистовство») — кульминация полового цикла, характеризующаяся готовностью самки к осеменению и оплодотворению. Метэструс — период затухания половой активности. Анэструс —период функционального покоя половой системы. А.П. Студенцов (1953), основываясь на поведенческих реакциях самки и морфофункциональных изменениях, выявляемых в репродуктивных органах клиническими методами, подразделил половой цикл на 3 стадии: возбуждения, торможения, уравновешивания. Такое подразделение полового цикла при очевидной односторонности (учитывается лишь динамика нервных процессов) получило широкое признание в нашей стране, поскольку дает четкие ориентиры относительно контроля половых циклов и выбора оптимального времени осеменения. Стадия возбуждения характеризуется последовательным вовлечением в цепь физиологических процессов течки, общей половой реакции, охоты, при наличии в яичниках созревающих фолликулов. Биологическое назначение течки состоит в подготовке половых путей к приему, сохранению и продвижению спермиев. Продуцируемая слизистыми покровами слизь обладает высокой абсорбционной способностью. Вместе с ней удаляются микробы и их токсины, погибшие лейкоциты, спермоантитела, десквамированный эпителий, что обеспечивает самоочищение половых путей. Вспомогательную роль выполняют фагоцитирующие лейкоциты, содержание которых в просвете маточных рогов возрастает в десятки раз. Наряду с этим формируется защитный барьер, препятствующий проникновению микроорганизмов в матку из внешней среды. Он включает ороговение и отторжение в просвет влагалища покровного эпителия, заполнение канала шейки матки вязкой слизью, усиление моторики матки в конце течки. Общая половая реакция характеризуется повышенной двигательной активностью самки, торможением молокоотдачи, подавлением пищевого рефлекса, прыжками на других самок. Значение этих поведенческих реакций состоит в том, что под влиянием нейросексуальных раздражителей, воспринимаемых органами зрения, обоняния и осязания, происходит выброс в кровь окситоцина, что сопровождается резким подъемом сократительной активности матки. В сочетании с принятием вертикальной позы при прыжках на других самок это обеспечивает интенсивную эвакуацию слизи. Охота является кульминационным звеном стадии возбуждения; это не что иное как строго дифференцированная реакции самки, проявляющаяся влечением к самцу данного вида животных и готовностью к садке и совокуплению. За проявления течки, общей половой реакции, охоты ответственны эстрогены, источниками которых служат созревающие фолликулы в яичниках самки. По мере роста и развития фолликулов суточная продукция эстрогенов непрерывно нарастает, достигая пика непосредственно перед началом охоты. Начало проявления и угасание каждого из феноменов стадии возбуждения полового цикла предопределяется пороговой чувствительностью к эстрогенам рецепторных зон соответствующих эффекторных органов. Стадия возбуждения, как правило, начинается с течки, поскольку рецепторы матки, влагалища, преддверия, половых губ имеют относительно низкий порог чувствительности к эстрогенам. На фоне нарастания признаков течки на нее наслаиваются общая половая реакция и затем охота. Зрелый фолликул разрывается, при этом яйцо попадает в яйцепровод, где становится доступным для спермиев. Стадия торможения характеризуется постепенным угасанием феноменов стадии возбуждения в последовательности, предопределенной пороговой чувствительностью контролирующих зон: вначале прекращается охота, затем половое возбуждение и в конце стадии торможения полностью исчезают признаки течки. Состояние яичников в стадию торможения характеризуется формированием желтых тел и началом становления их гормональной функции. Стадия уравновешивания — период относительной стабильности физиологических процессов в репродуктивных органах самки. В стадии уравновешивания отсутствуют признаки течки, общей половой реакции, сохраняется индифферентное отношение к самцу; в яичниках отсутствуют крупные полостные фолликулы. Функция яичников в этот период определяется наличием желтых тел, осуществляющих биосинтез прогестерона. Общая продолжительность полового цикла зависит от срока функционирования желтых тел. Половые циклы самок дифференцируют на полный и неполный. При полном половом цикле стадия возбуждения имеет яркие проявления течки, общей половой реакции, охоты и завершается овуляцией. Все это создает оптимальные условия для оплодотворения. Неполным называют половой цикл, при котором отдельные компоненты стадии возбуждения проявляются слабо либо полностью выпадают. При неполном половом цикле вероятность оплодотворения самки снижается. Помимо этого, различают ритмичные и аритмичные половые циклы. В первом случае длительность каждого последующего цикла соответствует предыдущему, во втором — циклы нормальной продолжительности чередуются с удлиненными либо укороченными половыми циклами. Неполноценность половых циклов может выражаться в увеличении или, наоборот, уменьшении продолжительности течки, охоты, задержке овуляции.
|
Последнее изменение этой страницы: 2019-05-06; Просмотров: 372; Нарушение авторского права страницы

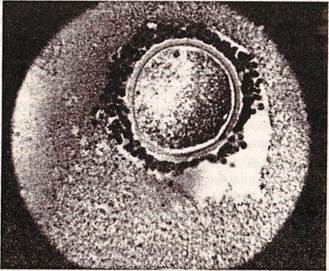
 Рис. 7. Третичный фолликул (граафов пузырёк): 1- яйцо; 2- прозрачная оболочка яйца; 3- лучистый венец; 4- зернистый слой(гранулёза); 5- яйценосный бугорок; 6- фолликулярная жидкость; 7- капсула (тека)
Рис. 7. Третичный фолликул (граафов пузырёк): 1- яйцо; 2- прозрачная оболочка яйца; 3- лучистый венец; 4- зернистый слой(гранулёза); 5- яйценосный бугорок; 6- фолликулярная жидкость; 7- капсула (тека)