|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Функциональная архитектура зрительной корыСтр 1 из 4Следующая ⇒
Функциональная архитектура зрительной коры
Функциональная архитектура зрительной коры
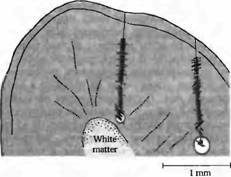
Зрительная кора организована в виде вертикальных кластеров клеток, имеющих сходные функциональные свойства. Нейроны, получающие информацию избирательно либо от правого, либо от левого глаза, сгруппированы в глазодоминантные колонки. Ориентационные колонки состоят из нейронов, которые имеют сходную чувствительность к углам ориентации линий и краев изображения. Глазодоминантные колонки и ориентационные колонки были впервые открыты при регистрации электрической активности от группы кортикальных клеток во время прохождения электродом вглубь зрительной коры. Глазодоминантные и ориентационные колонки могут быть также визуализованы при помощи биохимических и оптических методов, которые выявляют зоны активности коры у живого животного. Аксоны крупноклеточных (magnocellular. M) и мелкоклеточных (parvocellular, P) нейронов ядер латерального коленчатого тела проецируются в различные области 4 слоя первичной зрительной коры. В дальнейшем эта информация по М- или Р-трактам распространяется в различающиеся области первичной и вторичной зрительной коры. Нейроны М-пути отвечают за детекцию движущихся стимулов. Они также чувствительны к различиям в контрастности и интенсивности зрительных стимулов. Нейроны Р-пути ответственны за тонкие детали изображения и восприятие цвета. При восприятии изображения такие его элементы, как цвет и движение, анализируются независимо. Это иллюстрируется тем фактом, что повреждения в изолированных зонах мозга приводят к селективной потере одного из этих свойств, но не сказываются на общем восприятии изображения глазом. Повреждения в области париетальной коры, известной как МТ (или V5), приводят к полной потере способности определять движения объектов и к нарушениям в восприятии интенсивности изображения. При локализации повреждения в затылочно-височной области (зона V4) теряется способность к распознаванию цвета. Выдающимся достижением является использование неинвазивных методов функционального магнитного резонанса для определения зон активности мозга животных и человека. Хотя отдельные корковые глазодоминантные и ориентационные колонки имеют размеры меньше сегодняшнего предела разрешения данного метода, именно при помощи этого метода были найдены области зрительной коры, специализирующиеся на задачах особого рода, таких как определение движения и распознавание лиц. Путем анализа эффекта зрительных стимулов на последовательно связанные кортикальные клетки можно понять процессы переработки информации в первичной зрительной коре. Этот подход приблизил нас к пониманию клеточных механизмов анализа формы в каждой отдельной точке поля зрения. Сейчас же нашей задачей является изучить, каким образом в коре кодируются другие аспекты восприятия, такие как цвет и движение, и каким образом эти свойства могут быть затем собраны воедино при восприятии цельного изображения. Мы уже описали поточечное представление сетчатки в области зрительной зоны (V1) и то, каким образом происходит разделение информации от обоих глаз в колонках глазного доминирования. В пределах данной ретинотопической карты существует функциональное деление; так, например, определенные колонки клеток реагируют на линии строго определенной ориентации. Мы начинаем эту главу исследованием взаимосвязи между колонками глазного доминирования и ориентационными колонками, затем мы рассмотрим доказательства того, что в зрительной коре движение и цвет анализируются параллельно различными группами клеток. И, наконец, мы рассмотрим примеры высшего уровня обработки зрительной информации в зонах, расположенных вне первичной зрительной коры. Рис. 2. Выявление глазодоминантных колонок с помощью оптической регистрации. Чувствительная камера обнаруживает оптический сигнал в коре обезьяны во время стимуляции только одного глаза. Изменения в интенсивности сигнала кодируются при помощи цвета, поэтому активные зоны выглядят в виде светлых полос. Паттерн светлых полос соответствует колонкам глазного доминирования, выявляемым при маркировании анатомическими методами. Рис. 3. Оси ориентации рецептивных полей нейронов по мере погружения электрода вглубь коры мозга кошки. Все последующие клетки демонстрируют одинаковую ориентацию оси, что показано в виде угла, под которым располагается штриховая полоска по отношению к линии проникновения электрода. Погружение электрода справа идет с большим отклонением от перпендикуляра, следовательно, при пересечении им нескольких колонок часто меняется и ориентация оси. Расположение каждой клетки определяется при помощи нанесения небольших повреждений по мере погружения электрода и при завершении погружения (показано кружком). На основании этих данных производится реконструкция хода электрода. Подобные эксперименты позволили установить, что клетки со сходными свойствами у кошек и обезьян организованы в виде колонок, идущих под определенными углами к поверхности коры. Ориентационные колонки
Какие другие функциональные объединения возможны для зрительных нейронов в зоне V1? Ориентационные предпочтения простых и комплексных клеток. Таким образом, логичен вопрос: является ли данная особенность систематически повторяющейся в зрительной коре? Репрезентативный эксперимент, отвечающий на этот вопрос, представлен на рис. 3. Микроэлектрод погружается перпендикулярно поверхности вглубь серого вещества в зоне V1 мозга кошки. Каждый штрих на этом рисунке показывает расположение одной клетки и предпочтительную ориентацию ее рецептивного поля при предъявлении визуальных раздражителей в виде полосок. После погружения электрода в контрольной точке клетки локально повреждаются посредством пропускания электрического тока. Расположение каждой регистрируемой клетки затем реконструируется на основании этих контрольных точек (конечная точка показана кружком в конце пути электрода). Как показано с левой стороны рисунка, первые (начиная с поверхности) 38 клеток наиболее эффективно активировались полосками, расположенными под углом около 90 градусов к вертикальной оси. После погружения на глубину около 0, 6 мм ось ориентации рецептивного поля клеток сместилась примерно на 45 градусов. При другом погружении, показанном справа, с более острым углом к поверхности коры, каждая последующая клетка имеет разные варианты ориентации оси. При таком наклонном проникновении оси полей меняются по мере прохождения через разные колонки, имеющие отличающуюся ориентацию. Ориентационные колонки получают сигналы от клеток, рецептивные поля которых значительно перекрываются на поверхности сетчатки. Информация об организации ориентационных колонок в зрительной коре обезьян и кошек была впервые получена при проведении именно косых (тангенциальных), а не перпендикулярных погружений электрода в глубь коры. При продвижении электрода на каждые 50 мкм в горизонтальном направлении вдоль коры ориентация оси рецептивного поля меняется примерно на 10 градусов, иногда такие последовательные смены ориентации достигают в конечном итоге 180 градусов. Колонки ориентации (20-50 мкм) гораздо уже, чем глазодоминантные колонки (250-500 мкм). Первая анатомическая демонстрация ориентационных колонок, основанная на их разной функциональной активности, была произведена Соколовым, который использовал 2-деоксиглюкозу, хорошо захватывающуюся активными клетками. В результате метаболически активные клетки становились радиоактивными и их расположение можно было видеть на ауторадиограммах. У обезьян и кошек, которым показывали горизонтальные или вертикальные полосы, «полоски» радиоактивности совпадали с расположением колонок вертикальной или горизонтальной ориентации, различимых в коре без всякой окраски. Организация ориентационных колонок изучалась также при помощи оптических методов на живых животных. Примером может служить эксперимент Бонхоеффера и Гринвальда. Представление зрительных стимулов различной ориентации приводило к активности в различных кортикальных зонах. Ответы на разные ориентации показаны разными цветами. Удивительной является организация ориентационных колонок по отношению друг к другу. Сначала их организация кажется достаточно беспорядочной. Однако при тщательном исследовании можно заметить определенные центры в виде пестрого «волчка», в котором сходятся отдельные точки всех ориентаций. Отсюда клетки, ответственные за определенную ориентацию, расходятся в удивительно правильном порядке. Некоторые волчки ориентированы по часовой стрелке, некоторые — против. Таким образом, ориентация в коре представлена скорее в радиальном, чем в линейном виде. Каждый угол ориентации уникален (возникает только один раз) в пределах цикла, вертикальные и горизонтальные контуры занимают несколько большую площадь коры. На каждом квадратном миллиметре коры находятся один или два таких центра, равноудаленных друг от друга. Интересно то, что подобный способ организации коры был предложен ранее, на основании теоретических предпосылок. Рис. 4. «Пятна» в зоне V1 и «полоски» в зоне V2 зрительной коры обезьян, окрашенной на цитохромоксидазу. Пятна организованы в виде узора «в горошек». Можно различить четкую границу между зонами V1 и V2. По этой линии «пятна» переходят в «полоски», толстые и тонкие, идущие под определенным углом к границе. Рис. 5. Схематическая организация М-, Р- и К-кана-лов, направляющихся в зрительную кору. Функционально различные слои ЛКТ направляют отростки к различным слоям в V1. К-слои проецируются в «пятна» в слое 2 и 3. М- и Р-слои области 4С могут взаимодействовать предпочтительно с областями «пятна» и с областями, расположенными между пятен в слоях 2 и 3. «Пятна» проецируются преимущественно в тонкие полоски в V2. Тонкие полоски проецируются в V4. Толстые полосы в V2 получают информацию от слоя 4В в V и проецируются в ассоциативную зону МТ (V5). M-каналы проецируются в дорзальную (теменную) зрительную кору, где происходит анализ движения. Р-каналы проецируются предпочтительно в зону V4, где осуществляется обработка информации о цвете. Детекция движения и зона МТ
Если попросить дать краткое определение, что такое зрение, большинство из нас скажет о восприятии объектов, включающем анализ тонких деталей, примерно как это происходит во время чтения этих слов. Однако не менее важной функцией зрения является анализ движения, хотя эта функция и остается в основном непроизвольной и подсознательной (за исключением тех, кто из-за окуломоторного расстройства не может автоматически управлять перемещением зрительного поля). Как уже было описано, движение анализируется крупноклеточно-париетальной частью зрительного тракта (рис. 4). Нейроны крупноклеточного пути чувствительны к движущимся стимулам, это их свойство сохраняется в зонах V1 и V2 (в пределах толстых полос), через которые этот путь проходит, направляясь к средневисочной ассоциативной коре (зона МТ, или V5). Зона МТ также имеет ретинотопическое строение. Нейроны в этой области чувствительны к скорости и направлению движения стимула, а также объединены в группы в виде колонок со сходными предпочтительными направлениями движения. Когда небольшие области МТ химически повреждаются при помощи нейротоксина, у обезьяны нарушается способность воспринимать характер движения точек в соответствующих областях зрительного поля, хотя чувствительность к контрастности изображения при этом не меняется. Литература
1. Komatsu, H. 1998. Mechanisms of central color vision. Curr. Opin. Neurobiol. 8: 503-508. 2. Merigan, W. H., and Maunsell, J. H. R. 1993. How parallel are the primate visual pathways? Anna. Rev. Neurosci. 16: 369-402. 3. Newsome, W. T., and Wurlz, R. H. 1988. Probing visual cortical function with discrete chemical lesions. Trends Neunsci. 11: 394-400. 4. Tootell, R. В., Dale, A. M., Sereno, M. I., and Malach, R. 1996. New images from human visual cortex. Trends Neurosci. 19: 481-489. 5. Zeki, S. 1990. Colour vision and functional specialisation in the visual cortex. Disc. Neurosci. 6: 1-64. Функциональная архитектура зрительной коры
|
Последнее изменение этой страницы: 2019-10-04; Просмотров: 171; Нарушение авторского права страницы