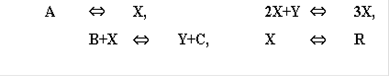|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Классические модели Лотки и Вольтерра ⇐ ПредыдущаяСтр 4 из 4
Первое понимание, что собственные ритмы возможны в богатой энергией системе за счет специфики взаимодействия ее компонентов пришло после появления простейших нелинейных моделей взаимодействия - химических веществ в уравнениях Лотки, и взаимодействия видов - в моделях Вольтерра [1, 3]. Уравнение Лотки рассмотрено им в 1926 г. в книге и описывает систему следующих химических реакций
В некотором объеме находится в избытке вещество А. Молекулы А с некоторой постоянной скоростью Запишем систему уравнений, описывающих реакцию:
Здесь X, Y, B - концентрации химических компонентов. Первые два уравнения этой системы не зависят от B, поэтому их можно рассматривать отдельно. При определенных значениях параметров в системе возможны затухающие колебания. Базовой моделью незатухающих колебаний служит классическое уравнение Вольтерра, описывающее взаимодействие видов типа хищник-жертва. Как и в моделях конкуренции (8), взаимодействие видов описывается в соответствии с принципами химической кинетики: скорость убыли количества жертв (x) и скорость прибыли количества хищников (y) считается пропорциональными их произведению
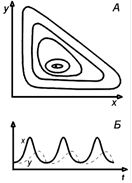
На рис. 6 представлены фазовый портрет системы, по осям которого отложены численности жертв и хищников - (а) и кинетика численности обоих видов - зависимость численности от времени-(б). Видно, что численности хищников и жертв колеблются в противофазе.
Рис.6. Модель хищник-жертва Вольтерра, описывающая незатухающие колебания численности. А. Фазовый портрет. Б. Зависимость численности жертвы и хищника от времени.
Модель Вольтерра имеет один существенный недостаток. Параметры колебаний ее переменных меняются при флуктуациях параметров и переменных системы. Такую систему называют негрубой. Этот недостаток устранен в более реалистичных моделях. Модификация модели Вольтерра с учетом ограниченности субстрата в форме Моно (уравнение 5) и учет самоограничения численности (как в уравнении 2) приводит к модели, подробно изученной А.Д. Базыкиным в книге " Биофизика взаимодействующих популяций" (1985).
Система (11 представляет собой некий кентавр, составленный из базовых уравнений (1, 2, 5, 10)и объединяющий их свойства. Действительно, при малых численностях и в отсутствие хищника жертва (x) будет размножаться по экспоненциальному закону(1). Хищник (y) в отсутствие жертв будут вымирать также по экспоненте. Если особей того или иного вида много, в соответствии с базовой моделью (2) срабатывает системный ферхюльстовский фактор (член -Ex2 в первом уравнении, и -My2 - во втором). Интенсивность взаимодействия видов считается пропорциональной произведению их численностей (как в модели (10)) и описывается в форме Моно (модель 5). Здесь роль субстрата играет вид-жертва, а роль микроорганизмов - вид-хищник. Таким образом, модель (11) брала в себя свойства базовых моделей (1), (2), (5), (10). Но модель (11) представляет собой не просто сумму свойств этих моделей. С ее помощью можно описать и гораздо более сложные типы поведения взаимодействующих видов: наличие двух устойчивых стационарных состояний, затухающие колебания численностей и проч. При некоторых значениях параметров система становится автоколебательной. В ней с течением времени устанавливается режим, при котором переменные изменяются периодически с постоянным периодом и амплитудой независимо от начальных условий. Волны жизни
До сих пор мы говорили о базовых моделях поведения живых систем во времени. Стремление к росту и размножению ведет к распространению в пространстве, занятию нового ареала, экспансии живых организмов. Жизнь распространяется так же как пламя по степи во время степного пожара. Эта метафора отражает тот факт, что пожар (в одномерном случае - распространение пламени по бикфордову шнуру) описывается с помощью той же базовой модели, что и распространение вида. Знаменитая в теории горения модель ПКП (Петровского - Колмогорова - Пискунова) впервые была предложена ими в 1937 г. именно в биологической постановке как модель распространения доминирующего вида в пространстве. Все три автора этой работы являются крупнейшими российскими математиками. Академик Иван Георгиевич Петровский (1901-1973) - автор фундаментальных трудов по теории дифференциальных уравнений, алгебре, геометрии, математической физике, в течение более 20 лет был ректором Московского Государственного университета им. М.В. Ломоносова. (1951-1973). Андрей Николаевич Колмогоров (1903 - ) глава российской математической школы по теории вероятностей и теории функций, автор фундаментальных трудов по математической логике, топологии, теории дифференциальных уравнений, теории информации, организатор школьного и университетского математического образования, написал несколько работ, в основу которых положены биологические постановки. В частности в 1936 г. он предложил и подробно исследовал обобщенную модель взаимодействия видов типа хищник-жертва (исправленный и дополненный вариант 1972).(См. Популяционная динамика) Рассмотрим постановку задачи о распространении вида в активной - богатой энергией (пищей) среде. Пусть в любой точке прямой r> 0 размножение вида описывается функцией f(x) = x(1-x). В начальный момент времени вся область слева от нуля занята видом x, концентрация которого близка к единице.. Справа от нуля - пустая территория. В момент времени t=0 вид начинает распространяться (диффундировать) вправо с константой диффузии D. Процесc описывается уравнением:
При t> 0 в такой системе начинает распространяться волна концентраций в область r> 0, которая является результатом двух процессов: случайного перемещения особей (диффузии частиц) и размножения, описываемого функцией f(x). С течением времени фронт волны перемещается вправо, причем его форма приближается к определенной предельной форме. Скорость перемещения волны определяется коэффициентом диффузии и формой функции f(x), и для функции f(x), равной нулю при x=0 и x=1 и положительной в промежуточных точках, выражается простой формулой: =2Df'(0). Изучение пространственного перемещения в модели хищник-жертва (10) показывает, что в такой системе в случае неограниченного пространства будут распространяться волны " бегства и погони", а в ограниченном пространстве установятся стационарные пространственные неоднородные структуры (диссипативные структуры), или авто волны, в зависимости от параметров системы. модель имитационный базовый математический Автоволны и диссипативные структуры. Базовая модель " брюсселятор"
На рассмотренной выше одномерной модели (14) видно, что взаимодействие нелинейной химической реакции и диффузии приводит к нетривиальным режимам. Еще более сложного поведения следует ожидать в двумерных моделях, описывающих взаимодействие компонентов системы. Первая такая модель была изучена Тьюрингом в работе под названием " Химические основы морфогенеза". Алан М. Тьюринг (1912-1954) английский математик и логик, прославился своими работами по компьютерной логике и теории автоматов. В 1952 г. он опубликовал первую часть исследования, посвященного математической теории образования структур в первоначально однородной системе, где одновременно проходят химические реакции, в том числе автокаталитические процессы, сопровождаемые потреблением энергии, и пассивные процессы переноса - диффузия. Это исследование осталось незаконченным, так как он покончил жизнь самоубийством, находясь под действием депрессантов, которыми его принудительно лечили в тюрьме, где он отбывал срок по обвинению в гомосексуализме. Работа Тьюринга стала классической, ее идеи легли в основу современной теории нелинейных систем, теории самоорганизации и синергетики. Рассматривается система уравнений:
Уравнения такого типа называются уравнениями " реакция-диффузия". В линейных системах диффузия процесс, который приводит к выравниванию концентраций во всем реакционном объеме. Однако в случае нелинейного взаимодействия переменных x и y, в системе может возникать неустойчивость гомогенного стационарного состояния и образуются сложные пространственно-временные режимы типа авто волн или диссипативных структур - стационарных во времени и неоднородных по пространству распределений концентраций, существование которых поддерживается в активных средах за счет потребления энергии системы в процессах диссипации. Условием возникновения структур в таких системах является различие коэффициентов диффузии реагентов, а именно, наличие близкодействующего " активатора" с малым коэффициентом диффузии и дальнодействующего " ингибитора" с большим коэффициентом диффузии. Такие режимы в двухкомпонентной системе были изучены в деталях на базовой модели под названием " брюсселятор" (Пригожин и Лефевр, 1968), названной в честь брюссельской научной школы под руководством И.Р. Пригожина, в которой наиболее интенсивно проводились эти исследования. Илья Романович Пригожин (род 1917 г. в Москве) - всю жизнь работал в Бельгии. С 1962 г. он - директор Международного Сольвеевского института физической химии в Брюсселе, а с 1967 г. - директор Центра статистической механики и термодинамики Техасского университета (США). В 1977 г. он получил Нобелевскую премию за работы по нелинейной термодинамике, в частности по теории диссипативных структур - устойчивых во времени неоднородных в пространстве структур. Пригожин является автором и соавтором целого ряда книг [" Термодинамическая теория структуры, устойчивости и флуктуаций", " Порядок из хаоса", " Стрела времени", и др.], в которых он развивает математические, физико-химические, биологические и философские идеи теории самоорганизации в нелинейных системах, исследует причины и закономерности рождения " порядка из хаоса" в богатых энергией открытых для потоков вещества и энергии системах, далеких от термодинамического равновесия, под действием случайных флуктуаций. Классическая модель " брюсселятор" имеет вид
и описывает гипотетическую схему химических реакций:
Ключевой является стадия превращения двух молекул x и одной молекулы y в x так называемая тримолекулярная реакция. Такая реакция возможна в процессах с участием ферментов с двумя каталитическими центрами. Нелинейность этой реакции в сочетании с процессами диффузии вещества и обеспечивает возможность пространственно-временных режимов, в том числе образование пространственных структур в первоначально однородной системе морфогенез. Заключение
Современная математическая биология использует различный математический аппарат для моделирования процессов в живых системах и формализации механизмов, лежащих в основе биологических процессов. Имитационные модели позволяют на компьютерах моделировать и прогнозировать процессы в нелинейных сложных системах, каковыми являются все живые системы, далекие от термодинамического равновесия. Базовые модели математической биологии в виде простых математических уравнений отражают самые главные качественные свойства живых систем: возможность роста и его ограниченность, способность к переключениям, колебательные и стохастические свойства, пространственно-временные неоднородности. На этих моделях изучаются принципиальные возможности пространственно-временной динамики поведения систем, их взаимодействия, изменения поведения систем при различных внешних воздействиях - случайных, периодических и т.п. Любая индивидуальная живая система требует глубокого и детального изучения, экспериментального наблюдения и построения своей собственной модели, сложность которой зависит от объекта и целей моделирования. Литература
1. Вольтерра В. Математическая теория борьбы за существование. М., Наука, 1976, 286 с. 2. Пайтген Х.-О., Рихтер П.Х. Красота фракталов. Образы комплексных динамических систем. М., Мир, 1993, 176 с. . Ризниченко Г.Ю., Рубин А.Б. Математические модели биологических продукционных процессов. М., Изд. МГУ, 1993, 301 с. . Романовский Ю.М., Степанова Н.В., Чернавский Д.С. Математическая биофизика. М., Наука, 1984, 304 с. . Рубин А.Б., Пытьева Н.Ф., Ризниченко Г.Ю. Кинетика биологических процессов. М., МГУ., 1988 . Свирежев Ю.М., Логофет. Устойчивость биологических сообществ М., Наука, 1978, 352c . Базыкин А.Д. Биофизика взаимодействующих популяций. М., Наука, 1985, 165 с. 8. J.D. Murray " Mathematical Biology", Springer, 1989, 1993. |
Последнее изменение этой страницы: 2020-02-16; Просмотров: 171; Нарушение авторского права страницы
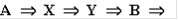
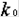 превращаются в молекулы вещества X (реакция нулевого порядка). Вещество X может превращаться в вещество Y, причем скорость этой реакции тем больше, чем больше концентрация вещества Y - реакция второго порядка. В схеме это отражено обратной стрелкой над символом y. Молекулы Y в свою очередь необратимо распадаются, в результате образуется вещество B (реакция первого порядка).
превращаются в молекулы вещества X (реакция нулевого порядка). Вещество X может превращаться в вещество Y, причем скорость этой реакции тем больше, чем больше концентрация вещества Y - реакция второго порядка. В схеме это отражено обратной стрелкой над символом y. Молекулы Y в свою очередь необратимо распадаются, в результате образуется вещество B (реакция первого порядка).

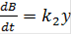 (9)
(9)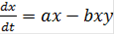
 (10)
(10)
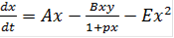
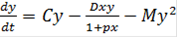 , (11)
, (11) (12)
(12)
 (13)
(13)
 (14)
(14)