|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ВИДЫ ВЗАИМОДЕЙСТВИЯ НЕАЛЛЕЛЬНЫХ ГЕНОВ
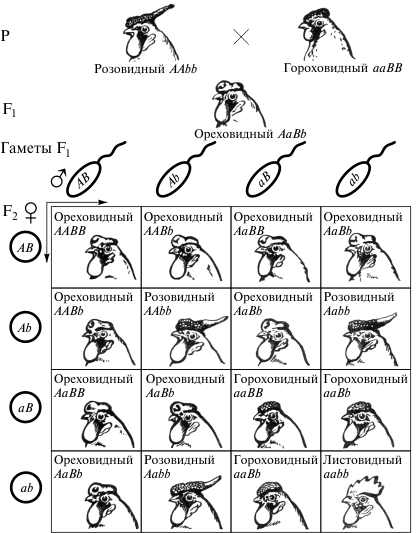
Выделяют три типа взаимодействия неаллельных генов: комплементарность, эпистаз и полимерия. Комплементарность – это вид взаимодействия неаллельных генов, когда при одновременном присутствии в генотипе особи доминантных аллелей разных генов формируется новый признак. Варианты расщеплений по фенотипу во втором поколении при комплементарном взаимодействи неаллелельных генов. Вариант расщепления 9: 3: 3: 1. Каждый доминантный аллель имеет свое проявление, тогда как два доминантных аллеля в сочетании формируют новый признак. Примером является формирования разных форм гребней у кур и петухов. Аллель P контролирует развитие гороховидного гребня у петухов (генотип P_rr), аллель R – розовидного (генотип ppR_), при отсутствии доминантных аллелей развивается простой (листовидный) гребень (генотип pprr), при наличии обоих доминантных аллелей формируется ореховидный гребень (генотип P_R_).
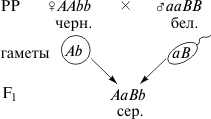
Вариант расщепления 9: 3: 4. Примером является наследование окраски шерсти у мышей (серый, черный, белый). Доминантный аллель (С)контролирует синтез черного пигмента, рецессивный аллель этого гена (с)не синтезирует черный пигмент (мыши белые), доминантный аллель (B) не аллельного гена обусловливает зональное распределение этого черного пигмента по волоску (на кончике и у основания волоска), поэтому при генотипе С_В_ – мыши серые, рецессивный аллель этого гена (b) обусловливает равномерное распределение пигмента по волоску, следовательно при генотипе С_bb мыши будут черными. При генотипах ссВ_ и ссbb мыши белые.
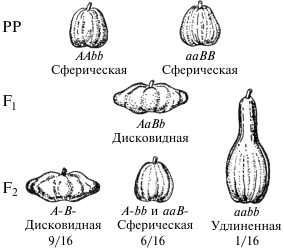
F2: 9 сер.: 3 черн.: 4 бел Вариант расщепления 9: 6: 1. При скрещивании двух разновидностей фигурной тыквы со сферической формой плода гибриды первого поколения обладают новым признаком — плоскими или дисковидными плодами. При скрещивании гибридов между собой в F2 наблюдается расщепление в соотношении 9 дисковидных: 6 сферических: 1 удлиненная. . 4. Вариант расщепления 9: 7. Комплементарное взаимодействие описано также при наследовании окраски цветов у душистого горошка. Большая часть сортов этого растения имеет пурпурные цветы с фиолетовыми крыльями, которые характерны для дикой сицилийской расы, но есть также сорта с белой окраской. Скрещивая растения с пурпурной окраской цветов с растениями с белыми цветами Бетсон и Пеннет установили, что пурпурная окраска цветов полностью доминирует над белой, и в F2 наблюдается соотношение 3: 1. Но в одном случае от скрещивания двух белых растений получилось потомство, состоящее только из растений с окрашенными цветами. При самоопылении растений F1 было получено потомство, состоящее из двух фенотипических классов: с окрашенными и неокрашенными цветами в соотношении 9/16: 7/16. Полученные результаты объясняются комплементарным взаимодействием двух пар неаллельных генов, доминантные аллели которых ( С и Р ) в отдельности не способны обеспечить развитие пурпурной окраски, так же как и их рецессивные аллели ( ссрр ). Окраска проявляется только при наличии в генотипе обоих доминантных генов, взаимодействие которых обеспечивает синтез пигмента. Однако такой же результат получается и в том случае, если взаимодействующие доминантные гены имеют одинаковое фенотипическое проявление. Например, формирование слуха у человека обусловлено наличием в генотипе доминантных аллелей разных генов (D – развитее улитки и E – развитие слухового нерва), а при отсутствии в генотипе одного из этих доминантных аллелей (генотипы D_ee иddE_) и при рецессивном гомозиготном генотипе (aabb) наблюдается глухонемота.
F2: 9 пурп.: 7 бел.
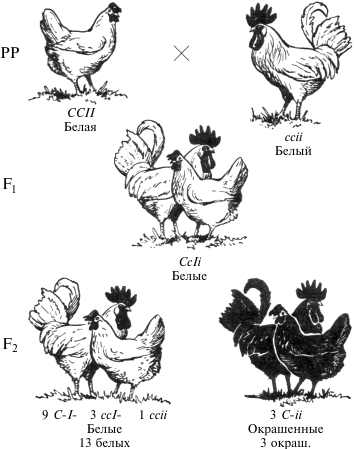
Эпистаз – это вид взаимодействия аллелей разных генов, характеризующийся подавлением проявления признака в присутствии супрессора. Различают доминантный и рецессивный эпистаз. При доминантном эпистазе доминантный аллель одного гена (эпистатического) подавляет проявление доминантного аллеля другого гена(гипостатического). Допустим, что при наличии аллеля А образуется фермент, разрушающий пигмент, синтезированный аллелем другого гена В. Тогда при сочетании А_В_ признак не формируется. Варианты расщеплений по фенотипу во втором поколении при доминантном эпистазе. 1. Вариант расщепления 13: 3. Примером является наследование и окраски оперения у кур. Доминантный аллель (С) гена детерминирует пигментацию оперения у кур, рецессивный аллель (с) не дает пигментацию. Доминантный аллель (I) неаллельного гена является супрессором, т.е.подавляет действие аллеля С, рецессивный аллель (i) – нейтральный. Поэтому куры с генотипами C_I_, ссI_ и ссii будут белыми, пигментация оперения наблюдается при генотипах C_ii.
2. Вариант расщепления 12: 3: 1. В качестве примера доминантного эпистаза можно рассмотреть наследование масти лошадей. Если C – серая масть (супрессор), В – черная, то лошади с генотипами C_B_, C_bb будут серыми, черная масть формируется при генотипах ссB_, при рецессивном гомозиготном генотипе (ссbb) лошади будут рыжими.
F2: 12 сер.: 3 черн.: 1 рыж.
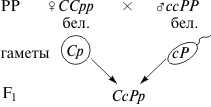
При рецессивном эпистазе рецессивные аллели одного гена, будучив гомозиготном состоянии, подавляют доминантный аллель другого гена. Примером рецессивного эпистаза у человека может служить, так называемый, бомбейский феномен, когда индивид, имеющий доминантный аллель группы крови системы АВО (например, IA или IB), идентифицируется как человек с I группой. Это обусловлено эпистатическим действием рецессивных аллелей hh аутосомного гена-супрессора, которые подавляют развитие антигенов групп крови. В этом случае, например, особи с генотипомIAI0hh будут иметь I группу крови. Отсутствие необходимых данных о роли первичных продуктов многих генов в формировании сложных признаков часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образованиях этих признаков. В одних случаях развитие признака при наличии двух аллелей разных генов в доминантном состоянии рассматривают как комплементарное взаимодействие, в других - отсутствие признака, определяемого одним из аллелей при отсутствии другого аллеля в доминантном состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантном эпистазе. С этой точки зрения разделение взаимодействия аллелей на комплементарное и эпистатическое несколько искусственно, ибо во всех этих случаях сложный признак является результатом сочетания в генотипе определенных аллелей соответствующих генов, которые обеспечивают синтез продуктов, участвующих в цепи биохимических преобразований на разных уровнях формирования сложного признака. Полимерия – это вид взаимодействия неаллельных генов, при котором доминантные аллели разных генов отвечают за проявления одного итого же признака. Полимерное взаимодействие может быть качественным – некумулятивная полимерия (наличие хотя бы одного доминантного аллеля приводит к формированию признака), или количественным – кумулятивная полимерия (степень проявления признака зависит от количества доминантных аллелей). По типу кумулятивной полимерии у человека наследуется интенсивность пигментации кожи: степень пигментации прямо пропорциональна количеству меланина и количеству доминантных аллелей. Четыре пары доминантных аллелей – генотип А1А1А2А2А3А3А4А4, обусловливают черный цвет кожи; четыре доминантных аллеля – темно-коричневый; три доминантных аллеля – коричневый; два и один доминантных аллеля – смуглый; отсутствие доминантных аллелей а1а1а2а2 а3а3а4а4 – светлая окраска кожи). Кумулятивная полимерия лежит в основе определения количественных признаков (рост, масса, возможно, уровень развития интеллекта). Расщепление по фенотипу при полимерном взаимодействии генов во втором поколении F2 дигибридного скрещивания выражается как 15: 1. Такой тип расщепления соответствует проявлению признака даже при наличии хотя бы одного доминантного аллеля (15 частей), тогда как при отсутствии доминантных аллелей признак не проявляется (1 часть с гомозиготным по рецессивным аллелям генотипом а1а1а2а2…). Особый вид представляет взаимодействие, обусловленное местом положения гена в системе генотипа - эффект положения. Непосредственное окружение, в котором находится ген, может сказываться на характере его экспрессии. Изменение активности гена, наблюдаемое при хромосомных перестройках, нередко связано с перемещением его в другую группу сцепления при транслокациях или изменением его положения в своей хромосоме при инверсиях. Особый случай, очевидно, представляет изменение экспрессии генов в результате деятельности подвижных генетических элементов, активирующих или угнетающих проявление генов, вблизи которых они встраиваются. Наконец, большое значение в объединении генов в единую систему генотипа имеют регуляторные взаимодействия, обеспечивающие регуляцию генной активности. Продукты генов-регуляторов - белки-регуляторы обладают способностью узнавать определенные последовательности ДНК, соединяться с ними, обеспечивая, таким образом, транскрибирование информации со структурных генов или препятствуя транскрипции.
|
Последнее изменение этой страницы: 2020-02-16; Просмотров: 73; Нарушение авторского права страницы