|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Генов при научении происходит
Экспрессия «поздних» генов
Как уже упоминалось, многие «ранние» гены кодируют транскрипционные факторы, регулирующие активность наборов эффекторных «поздних» генов.(Поэтому, если долговременные изменения в мозге при формировании памяти действительно инициируются продуктами «ранних» генов, то геномный ответ нервных клеток на обучение должен быть таким же, как и у других клеток на факторы роста — т. е. двухфазным: вначале должна происходить экспрессия ранних генов, а затем активация регулируемых ими генов-мишеней. Еще в 1969 г. предположение о подобной двухстадийной активации генома при научении высказал Э. Глассман: «Можно предположить следующую последовательность химических событий, которые ведут к консолидации долговременной памяти: белок-1 ® РНК ® белок-2... Образование белка-1 основывается на сообщении о том, что у золотых рыбок пуромицин одновременно предотвращает изменения синтеза РНК и нарушает память. Если окажется, что правильны другие объяснения этого феномена, то данный белок можно будет с легкостью исключить из схемы. Пока же соблазнительно предположить, что этот белок является активатором специфических генов, которые кодируют РНК, образующуюся на следующем этане» (Classman, 1969, с. 635). И действительно, при стимуляции нервной системы транскрипционные факторы, кодируемые «ранними» генами, инициируют «вторую волну» синтеза белка, которая начинается через несколько часов после первоначального воздействия (Анохин, 1997). Так, при исследовании динамики синтеза РНК и белка в гиппокампе крыс после обучения активному избеганию было обнаружено, что увеличение включения радиоактивных предшественников возрастает в течение первого часа после сеанса, потом падает и приблизительно 3 часа находится на контрольном уровне, а затем, на 6-10 часу, вновь существенно возрастает (РоЫе, Matthies, 1974; Ruthrich et al., 1974; Popov et al., 1976). В полном соответствии с этим обстоятельством, введение в мозг ингибитора синтеза белка анизомицина через 3-5 часов после обучения нарушает консолидацию долговременной памяти (Tiunova et al., 1998). При этом сами гены транскрипционных факторов, таких как с-fos и c-jun, во время второй фазы синтеза РНК не экспрессируются (Анохин, 1997). Какие же эффекторные гены активируются в мозге под воздействием индуцируемых транскрипционных факторов?
В составе второй волны экспрессии генов при Научении активируются морфорегуляторные Молекулы
Среди «поздних генов» лучше всего исследованы мишени транскрипционных белков fos/jun, имеющие участки связывания с АР-1 элементами ДНК. АР-1 элемент присутствует в промоторных областях большого числа генов, многие из которых активируются в ответ на разнообразные экстраклеточные воздействия (Sheng, Greenberg, 1990). В число генов, содержащих АР-1 участок, входят, например, гены препроэнкефалина, S-100, нейрофиламентов, тирозингидроксилазы и N-CAM (Prentice et al., 1987; Lindenbaum et al, 1988; Masiakowski, Shooter, 1988; Sonnenberg et al., 1989; Sheng, Greenberg, 1990). Гены молекул N-CAM (neural cell adhesion molecules), относящиеся к семейству генов молекул клеточной адгезии, представляют в этом отношении особый интерес. Молекулы клеточной адгезии, или «морфорегуляторные молекулы» (Edelman, 1988), экспрессируясь на поверхности клеточных мембран, регулируют агрегацию и дисагрегацию клеток в процессах развития (Edelman, Jones, 1995). Блокада экспрессии или связывания молекул клеточной адгезии ведет к нарушениям морфогенетических паттернов в развитии. Ген N-САМ экспрессируется как в эмбриональном, так и во взрослом мозге. Мыши с направленной мутацией гена N-CAM имеют измененнную морфологию мозга, нарушения поведения и обучения (Tomasievich et al., 1993; Cremer et al., 1995). Особенно интересно, что функционально активные молекулы клеточной адгезии образуются во время второй волны синтеза белка после обучения (Scholey et al., 1993, Mileusnic et al., 1995). Антитела к молекулам клеточной адгезии способны вызвать у животных амнезию только при введении в течение строго фиксированного критического периода после обучения. Этот период охватывает интервал от 6 до 8 часов после обучения у крыс (Doyle et al., 1992) и от 4 до 6 часов у цыплят (Mileusnic et al., 1995; Scholey et al., 1995), и совпадает со временем, когда после обучения должны активироваться гены-мишени для продуктов ранних генов. В совокупности с тем, что гены N-CAM несут в своих промоторах АР-1 элементы, связывающиеся с транскрипционными факторами семейства fos/jun, это дает основания полагать, что они включаются при обучении в каскад молекулярных событий, индуцируемых «ранними» генами. В результате реактивации во взрослом мозге этих и других контролирующих развитие морфорегуляторных молекул нервные клетки могут прибретать при формировании нового опыта способность к перестройке своих синаптических контактов и специализации относительно вновь образующихся функциональных систем (Rose, 1995; Анохин, 1996, 1997).
Генетическом уровне научение Составляет с развитием Единый континуум
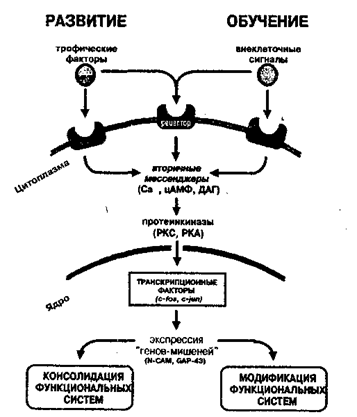
Рис.20.5. Общность молекулярных механизмов регуляции экспрессии генов при развитии нервной системы и научении В определенном смысле мы можем сказать, что на молекулярном уровне научение выступает как непрекращающийся процесс развития (Анохин, 1996).Однако механизмы регуляции экспрессии генов при научении имеют одно чрезвычайно важное отличие от сходных процессов в развитии.
Популярное:
|
Последнее изменение этой страницы: 2016-03-26; Просмотров: 643; Нарушение авторского права страницы
 20.5. На молекулярно-
20.5. На молекулярно-  Таким образом, при научении в нервных клетках наблюдается следующая последовательность молекулярно-генетических процессов. Вначале рассогласование текущей ситуации с имеющимся опытом запускает активацию каскада «ранних» регуляторных генов в группах клеток, опосредующих эти процессы. Продукты «ранних» генов индуцируют, в свою очередь, экспрессию «поздних» генов, в том числе генов морфорегуляторных молекул, являющихся ключевыми участниками процессов морфогенеза при эмбриональном развитии. Эти и другие эффекторные гены стабилизируют участие нейронов в новой, сложившейся в результата обучения, функциональной системе. При этом основные молекулярно-генетические элементы и этапы молекулярного каскада дифференцировки клетки оказываются чрезвычайно сходными при научении и развитии (рис. 20.5).
Таким образом, при научении в нервных клетках наблюдается следующая последовательность молекулярно-генетических процессов. Вначале рассогласование текущей ситуации с имеющимся опытом запускает активацию каскада «ранних» регуляторных генов в группах клеток, опосредующих эти процессы. Продукты «ранних» генов индуцируют, в свою очередь, экспрессию «поздних» генов, в том числе генов морфорегуляторных молекул, являющихся ключевыми участниками процессов морфогенеза при эмбриональном развитии. Эти и другие эффекторные гены стабилизируют участие нейронов в новой, сложившейся в результата обучения, функциональной системе. При этом основные молекулярно-генетические элементы и этапы молекулярного каскада дифференцировки клетки оказываются чрезвычайно сходными при научении и развитии (рис. 20.5).