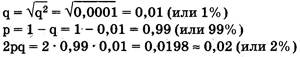|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
История изучения нуклеиновых кислотСтр 1 из 3Следующая ⇒
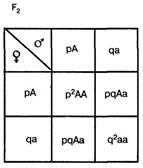
Закон Харди- Вайнберга Этот закон утверждает, что частоты аллелей и генотипов в данной популяции будут оставаться постоянными из поколения в поколение при выполнении следующих условий: 1) численность особей популяции достаточно велика (в идеале — бесконечно велика) 2) спаривание происходит случайным образом (т. е. осуществляется панмиксия), 3) мутационный процесс отсутствует, 4) отсутствует обмен генами с другими популяциями, 5) естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны. Иногда этот закон формулируют иначе: в идеальной популяции частоты аллелей и генотипов постоянны. Математическая модель закона отвечает формуле: p2+2pq+q2=1 Она выводится на основе следующих рассуждений. В качестве примера возьмем простейший случай — распределение двух аллелей одного гена. Пусть два организма являются основателями новой популяции. Один из них является (АА), а другой — (аа). Естественно, что все их потомство в F1 будет единообразным и будет иметь генотип (Аа). Далее особи F1 будут скрещиваться между собой. Обозначим частоту встречаемости доминантного аллеля (А) буквой p, а рецессивного аллеля (а) — буквой q. Поскольку ген представлен всего двумя аллелями, то сумма их частот равна единице, т. е. р + q = 1. Рассмотрим все яйцеклетки в данной популяции. Доля яйцеклеток, несущих доминантный аллель (А), будет соответствовать частоте этого аллеля в популяции и, следовательно, будет составлять р. Доля яйцеклеток, несущих рецессивный аллель (а), будет соответствовать его частоте и составлять q. Проведя аналогичные рассуждения для всех сперматозоидов популяции, придем к заключению о том, что доля сперматозоидов, несущих аллель (А), будет составлять р, а несущих рецессивный аллель (а) — q. Теперь составим решетку Пеннета, при этом при написании типов гамет будем учитывать не только геномы этих гамет, но и частоты несомых ими аллелей. На пересечении строк и столбцов решетки мы получим генотипы потомков с коэффициентами, соответствующими частотам встречаемости этих генотипов.
Из приведенной решетки видно, что в F2 частота доминантных гомозигот (АА) составляет р, частота гетерозигот (Аа) — 2pq, а рецессивных гомозигот (аа) — q. Поскольку приведенные генотипы представляют собой все возможные варианты генотипов для рассматриваемого нами случая, то сумма их частот должна равняться единице, т. е. p2+2pq+q2=1 Главное применение закона Харди—Вайнберга в генетике природных популяций — вычисление частот аллелей и генотипов. Рассмотрим пример использования этого закона в генетических расчетах. Известно, что один человек из 10 тыс. является альбиносом, при этом признак альбинизма у человека определяется одним рецессивным геном. Давайте вычислим, какова доля скрытых носителей этого признака в человеческой популяции. Если один человек из 10 тыс. является альбиносом, то это значит, что частота рецессивных гомозигот составляет 0, 0001, т. е. q2 = 0, 0001. Зная это, можно определить частоту аллеля альбинизма q, частоту доминантного аллеля нормальной пигментации р и частоту гетерозиготного генотипа (2pq). Люди с таким генотипом как раз и будут скрытыми носителями альбинизма, несмотря на то что фенотипически этот ген не будет у них проявляться и они будут иметь нормальную пигментацию кожи.
Из приведенных простых расчетов видно, что, хотя число альбиносов крайне невелико — всего лишь один человек на 10 тыс., ген альбинизма несет значительное количество людей — около 2%. Иными словами, даже если признак фенотипически проявляется очень редко, то в популяции присутствует значительное количество носителей этого признака, т. е. особей, имеющих этот ген в гетерозиготе.
Благодаря открытию закона Харди—Вайнберга процесс микроэволюции стал доступен непосредственному изучению: о его ходе можно судить по изменениям из поколения в поколение частот генов (или генотипов). Таким образом, несмотря на то что этот закон действителен для идеальной популяции, которой нет и не может быть в природе, он имеет огромное практическое значение, так как дает возможность рассчитать частоты генов, изменяющиеся под влиянием различных факторов микроэволюции. Кариотип человека Кариотип- хромосомный комплекс вида со всеми его особенностями: числом хромосом, их формой, деталями строения. Кариотип человека содержит 46 хромосом (23 пары). 22 пары называются аутосомами, 23-я пара – половыми хромосомами. Метафазная хромосома состоит из двух хроматид, соединенных центромерой. Центромера делит хромосому на два плеча – длинное (q) и короткое (р).
В кариотипе человека различают следующие типы хромосом: 1. Метацентрические (равноплечие). У метацентрических хромосом плечо р равно по длине плечу q. 2. Субметацентрические (неравноплечие). У субметацентрических хромосом плечо р короче плеча q. 3. Акроцентрические. У акроцентрических хромосом плечо р имеет спутники, представляющие собой районы ядрышкового организатора. Здесь находятся гены рРНК. Спутники соединяются с коротким плечом спутничной нитью.
По морфологическим признакам и размерам хромосомы в кариотипе человека делятся на 7 групп. Группа А (1- 3 пары) – большие метацентрические хромосомы; Группа В (4, 5 пары) – большие субметацентрические, различаются только при специальном (дифференциальном окрашивании); Группа С (6 – 12 пары) – средние субметацентрические, различаются при дифференциальном окрашивании; Группа D (13 -15 пары) – большие акроцентрические, различаются при дифференциальном окрашивании; Группа Е (16 – 18 пары) – средние метацентрические (16) и субметацентрические (17, 18), 17 и 18 пары хорошо различимы при дифференциальном окрашивании. Группа F (19 и 20 пары) – маленькие метацентрические хромосомы; различаются при дифференциальном окрашивании; Группа G (21 и 22 пары) – маленькие акроцентрические, различаются при дифференциальном окрашивании; Половые хромосмы (23-я пара). У женщин это две Х-хромосомы, у мужчин – Х и У хромосомы. Х-хромосома при стандартном окрашивании неотличима от аутосом группы С. У-хромосому дифференцируют от хромосом группы G (она не имеет спутников, длина короткого плеча изменчива и наследуется от отца к сыну). У-хромосому можно выявить специфическим окрашиванием акрихин-ипритом. При таком окрашивании концевой участок длинного плеча выявляется в виде яркого пятна диаметром 0, 3-1, 0 мкм. Методи антропогенетики. Генеалогический метод позволяет преодолеть сложности, возникающие в связи с невозможностью скрещивания и малоплодностью человека. Если есть родословные, то можно, используя суммарные данные по нескольким семьям определить тип наследования (доминантный, рецессивный, сцеплённый с полом, аутосомный) признака, а также его моногенность или полигенность. Так, доминантный признак «габсбургская губа» (толстая выпяченная нижняя губа) прослеживается в династии Габсбургов, начиная с XV в. Аналогичное наследование легко выявляется для признака брахидактилия или короткопалость, вследствие недоразвития (срастания) концевых фаланг. По доминантному типу наследуя такой дефект, как ахондроплазия – карликовость, связанная с резким укорочением конечностей и др. Близнецовый метод используется для выяснения степени наследственной обусловленности исследуемых признаков. Явление полиэмбрионии известно у некоторых животных. Оно характеризуется появлением нескольких идентичных, или однояйцовых близнецов (ОБ) – многозиготных близнецов. Наряду с такими ОБ существуют разнояйцовые близнецы (РБ), рождающиеся при оплодотворении одновременно созревающих яйцеклеток. Если ОБ как результат кланового размножения одной оплодотворённой яйцеклетки всегда идентичны по полу и очень похожи, часто практически неразличимы, то РБ могут иметь как одинаковый, так и разный пол. Встречаются РБ, сильно различающиеся по внешним признакам, как различаются особи, возникшие в результате самостоятельных случаев оплодотворения. В этом случае РБ представляют результат расщепления при скрещивании Близнецовый метод основан на трёх положениях: 1. ОБ имеют идентичные генотипы, а РБ различные генотипы. 2. Среда, в которой развиваются близнецы и под действием которой появляются различия признаков у ОБ, может быть одинаковой и неодинаковой для одной и той же пары ОБ. 3. Все свойства организма определяются взаимодействием только двух факторов: генотипа и среды. ОБ и РБ обычно сравнивают по ряду показателей на большом материале. На основе полученных данных вычисляют показатели конкордантности (частоты сходства) и дискордантности (частоты различий).
Цитогенетический метод. Довольно большое число трудно отличимых друг от друга (в пределах групп) хромосом создавали трудности в применении цитологического метода и в развитии цитогенетики человека. Разработка методов дифференциальной окраски упростила проблему идентификации всех хромосом человека. Благодаря культивированию клеток человека в vitro можно получать достаточно большой материал для описания цитологических особенностей исследуемого индивидуума. Для этого обычно используют кратковременную культуру лейкоцитов периферической крови. Цитологический метод приобрёл большое значение в связи с возможностями, которые открыла гибридизация соматических клеток. Получение гибридов между соматическими клетками человека и мыши позволяет в значительной степени преодолеть проблемы, связанные с невозможностью скрещиваний и картировать многие гены, контролирующие метаболизм клетки. Популяционный метод, или методы генетики популяций широко применяются в исследованиях человека. Он даёт информацию о степени гетерозиготности и полиморфизма человеческих популяций, выявляет различия частот аллей между разными популяциями. Так, хорошо изучено распространение аллей системы групп крови АВО. Различную концентрацию конкретных аллей локуса 1 связывают с известными данными о чувствительности разных генотипов к инфекционным болезням. Это помогает понять направление эволюции и отбора, действовавшего в разных регионах, в истории человечества. Популяционный метод позволяет определить адаптивную ценность конкретных генотипов. Многие признаки и соответственно обусловливающие их гены адаптивно нейтральны и проявляются как естественный полиморфизм человеческих популяций (например, многие морфологические признаки: цвет глаз, волос, форма ушей и т.д.). Другие признаки возникли как адаптивные по отношению к определенным условиям существования; например, темная пигментация кожи негров предохраняет от действия солнечной радиации. Известны примеры условно адаптивных аллелей. К их числу относится такая генетическая аномалия, как серповидноклеточная анемия. Рецессивная аллель, вызывающая в гомозиготном состоянии это наследственное заболевание, выражается в замене всего одного аминокислотного остатка? -цепи молекулы гемоглобина. В популяциях человека так же, как и в популяциях других организмов, в гетерозиготном состоянии содержится значительный генетический груз, т. е. рецессивные аллели, приводящие к развитию различных наследственных болезней. Повышение степени инбридинга в популяциях должно приводить к повышению частоты гомозиготации рецессивных аллелей. Эта закономерность должна предостерегать от заключения близкородственных браков. онтогенетический метод, согласно которому развитие нормальных и патологических признаков рассматривается в ходе индивидуального развития. Методы учёта мутаций 1. метод СІВ. Наиболее объективно можно учитывать частоту возникновения рецессивных летальных мутаций, приводящих в гомозиготном состоянии к смерти несущих их особей. Генетическая структура характеризуется тем, что одна их Х-хромосом маркирована доминантным геном В и инверсией С. Эта инверсия препятствует кроссинговеру и обладает рецессивным летальным эффектом. 2. Для учёта рецессивных летальных мутаций, сцепленных с полом, применяют метод Мёллер-5. Самки линии м-5 несут в своих хромосомах по 2 инверсии в обеих Х-хромосомах: sc3 и δ 49. В этой системе кроссинговер полностью подавлен. Используемые инверсии не имеют рецессивного летального действия. Кроме того, обе хромосомы М-5 несут три маркера: два рецессивных- абрикосовый цвет глаз и укороченные щетинки и один доминантный Bar. При скрещивании исследуемых самцов с самками М-5 в индивидуальных семьях получают по два класса самок и самцов, если в Х-хромосоме сперматозоидов исходного самца не возникла рецессивная летальная мутация, то в соответствующей индивидуальной культуре F2 мы получим только один класс самцов. Этот метод можно использовать т для учёта мутаций Х-хромосом с видимым проявлением. Для этого применяется метод дабл-еллоу. 3. Для изучения рецессивных летальных мутаций, а также для рецессивных мутаций с видимым проявлением исследуемых мух скрещивают с мухами Cyl/Pm. В F1 получают гетерозигот по той или иной хромосоме исследуемой линии, и индивидуально скрещивают сегрегантов Cyl с мухами Cyl/Pm. В F2 и6ндивидуальных культур скрещивают между собой самцов и самок с признаками Cyl и анализируют F3. В отсутствие рецессивной летальной мутации расщепление будет 2 CyL: 1 Cy+ L+., а если в половых клатках мух исходной линии возникли летальные мутации, то в соответствующих индивидуальных культурах в F3 не будет нормальных мух. Физические мутагены К физическим мутагенам относятся: ионизирующее излучение (альфа-, бета-, гамма-, нейтронное и рентгеновское излучение), коротковолновое ультрафиолетовое излучение, СВЧ-излучение, действие экстремальных температур. Действие ионизирующего излучения основано на ионизации компонентов цитоплазмы и ядерного матрикса. При ионизации возникают высокоактивные химические вещества (например, свободные радикалы), которые различным образом действуют на клеточные структуры. Рассмотрим наиболее изученные механизмы мутагенного воздействия ионизирующего излучения. 1. Непосредственное воздействие частиц с высокой энергией на ДНК, которое приводит к ее разрывам: одиночным (под воздействием гамма-квантов, рентгеновских лучей) или множественных (под воздействием альфа-частиц, нейтронного излучения). Это универсальный механизм возникновения хромосомных перестроек на всех стадиях клеточного цикла, но он действует очень грубо – обычно клетки теряют способность к нормальному делению и погибают. К разрывам ДНК приводит и ультрафиолетовое облучение. 2. Опосредованное воздействие ионизирующих факторов связано с нарушением структуры ферментов, контролирующих репликацию, репарацию и рекомбинацию ДНК. Этот механизм наиболее эффективно действует на синтетической стадии интерфазы. При больших дозах мутагенов клетки погибают. (Поскольку раковые клетки делятся непрерывно, то облучение является универсальным средством подавления развития метастазов при онкологических заболеваниях – непрерывно делящиеся раковые клетки более уязвимы, чем медленно пролиферирующие или непролиферирующие нормальные клетки.) Опосредованное воздействие ионизирующих факторов индуцирует самые разнообразные генные и хромосомные мутации. При опосредованном действии ионизирующих факторов их мутагенный эффект может быть снижен с помощью специальных веществ – радиопротекторов. К радиопротекторам относятся различные антиоксиданты, взаимодействующие с продуктами ионизации. В то же время, мутагенный эффект может быть усилен, например, высокая температура повышает мутагенный эффект радиации. 3. Особенности мутагенного действия ультрафиолетовых лучей. ДНК интенсивно поглощает жесткий ультрафиолет с длиной волны ≈ 254 нм. Основным продуктом является образование нуклеотидных димеров: два нуклеотида, расположенных рядом в одной цепи ДНК, «замыкаются» сами на себя, образуя пары «тимин–тимин» или «тимин–цитозин». При репликации ДНК напротив такой пары в достраивающейся цепи могут стать два любых нуклеотида, т.е. принцип комплементарности не выполняется. Ультрафиолетовый свет – это сравнительно мягкий мутаген, поэтому его широко используют в селекции растений, облучая проростки. 4. Особенности мутагенного действия экстремальных температур. Собственный мутагенный эффект экстремальных температур не доказан. Однако очень низкие или очень высокие температуры нарушают деление клетки (возникают геномные мутации). Экстремальные температуры усиливают действие других мутагенов, поскольку снижают ферментативную активность репарационных систем. Химические мутагены К химическим мутагенам относятся самые разнообразные вещества. Рассмотрим мутагенное действие некоторых из них. Алкилирующие агенты. Вызывают алкилирование ДНК (например, метилирование, этилирование и т.д.). В результате при репликации ДНК нарушается принцип комплементарности, и происходит замена нуклеотидных пар: ГЦ → АТ; ГЦ → ЦГ; ГЦ → ТА Некоторые из алкилирующих агентов в природе не встречаются, их не распознают ферменты защитных систем. Такие вещества называются супермутагенами (например, N-метил-N-нитрозомочевина). Супермутагены применяются в селекции растений для получения индуцированных мутаций; их используют также как стимуляторы роста (в сверхмалых концентрациях). Гидроксиламин. Избирательно аминирует цитозин, что также нарушает принцип комплементарности при репликации ДНК. В результате происходит замена ГЦ → АТ. Нитриты. Осуществляют окислительное дезаминирование гуанина, аденина, цитозина. Также нарушается принцип комплементарности при репликации ДНК. В результате происходит замена АТ → ГЦ. Аналоги оснований. Это вещества, сходные с «обычными» азотистыми основаниями. Однако они способны образовывать комплементарные пары с разными «нормальными» основаниями. Например, при репликации ДНК напротив гуанина вместо цитозина достраивается 5-бромурацил (аналог тимина). В дальнейшем напротив 5-бромурацила достраивается аденин, а напротив аденина – обычный тимин. Этот же процесс может идти и в противоположную сторону. В результате происходят замены: ГЦ → АТ или АТ → ГЦ. Существует множество иных химических факторов, обладающих мутагенным, канцерогенным и тератогенным действием. Например, ионы тяжелых металлов, связываясь с ферментами репликации, репарации и рекомбинации, снижают их ферментативную активность. Таким образом, не являясь собственно мутагенами, ионы тяжелых металлов способствуют появлению мутаций. Кроме того, нужно учесть, что воздействие совершенно разных мутагенов может приводить к сходным результатам. Установлено, что мутагены при определенных условиях оказывают канцерогенное и тератогенное действие. Канцерогены – это факторы, провоцирующие развитие онкологических заболеваний; тератогены – это факторы, провоцирующие развитие различных аномалий, уродств. Тератогенный эффект дают многие лекарственные препараты. Например, в 1960-е гг. на Западе широко использовалось снотворное талидомид, применение которого привело к рождению большого числа детей с недоразвитыми конечностями. Наряду с тератами – уродствами – часто встречаются морфозы – изменения, которые не ведут к утрате органом его функций. Отличить мутагенное действие от тератогенного сравнительно легко: тераты (уродства) являются ненаследственными модификациями, они предсказуемы (направлены) и не сохраняются в последующих поколениях. Организация генома эукариот ПЛАСТИДНАЯ НАСЛЕДСТВЕННОСТЬ Во многих случаях изменения в структуре и функциях пластид связаны с мутациями одного хромосомного гена. У кукурузы, ячменя и некоторых других культур изучены многочисленные хлорофильные мутации, наследующиеся по правилам Г. Менделя. Однако часто наследование таких изменений не подчиняются менделеевским закономерностям, и объяснить его можно только исходя из представления о генетической непрерывности пластид. У ночной красавицы имеется пестролистная разновидность. На одном и том же растении наряду с зелеными ветвями имеются ветви с листьями, на которых зеленая ткань чередуется с бесцветными полосами и пятнами. Цветки на зеленых ветвях такого пестролистного растения независимо от того, какой пыльцой опылять их, дают семена, из которых всегда вырастают нормальные зеленые растения. Семена с ветвей, листья на которых лишены зеленой окраски, дают неокрашенные бесхлорофилльные проростки. Из семян, завязавшихся на пестролистных побегах, образуется смешанное в различном соотношении потомство, состоящее из зеленых, пестролистных и неокрашенных растений Закон Харди- Вайнберга Этот закон утверждает, что частоты аллелей и генотипов в данной популяции будут оставаться постоянными из поколения в поколение при выполнении следующих условий: 1) численность особей популяции достаточно велика (в идеале — бесконечно велика) 2) спаривание происходит случайным образом (т. е. осуществляется панмиксия), 3) мутационный процесс отсутствует, 4) отсутствует обмен генами с другими популяциями, 5) естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны. Иногда этот закон формулируют иначе: в идеальной популяции частоты аллелей и генотипов постоянны. Математическая модель закона отвечает формуле: p2+2pq+q2=1 Она выводится на основе следующих рассуждений. В качестве примера возьмем простейший случай — распределение двух аллелей одного гена. Пусть два организма являются основателями новой популяции. Один из них является (АА), а другой — (аа). Естественно, что все их потомство в F1 будет единообразным и будет иметь генотип (Аа). Далее особи F1 будут скрещиваться между собой. Обозначим частоту встречаемости доминантного аллеля (А) буквой p, а рецессивного аллеля (а) — буквой q. Поскольку ген представлен всего двумя аллелями, то сумма их частот равна единице, т. е. р + q = 1. Рассмотрим все яйцеклетки в данной популяции. Доля яйцеклеток, несущих доминантный аллель (А), будет соответствовать частоте этого аллеля в популяции и, следовательно, будет составлять р. Доля яйцеклеток, несущих рецессивный аллель (а), будет соответствовать его частоте и составлять q. Проведя аналогичные рассуждения для всех сперматозоидов популяции, придем к заключению о том, что доля сперматозоидов, несущих аллель (А), будет составлять р, а несущих рецессивный аллель (а) — q. Теперь составим решетку Пеннета, при этом при написании типов гамет будем учитывать не только геномы этих гамет, но и частоты несомых ими аллелей. На пересечении строк и столбцов решетки мы получим генотипы потомков с коэффициентами, соответствующими частотам встречаемости этих генотипов.
Из приведенной решетки видно, что в F2 частота доминантных гомозигот (АА) составляет р, частота гетерозигот (Аа) — 2pq, а рецессивных гомозигот (аа) — q. Поскольку приведенные генотипы представляют собой все возможные варианты генотипов для рассматриваемого нами случая, то сумма их частот должна равняться единице, т. е. p2+2pq+q2=1 Главное применение закона Харди—Вайнберга в генетике природных популяций — вычисление частот аллелей и генотипов. Рассмотрим пример использования этого закона в генетических расчетах. Известно, что один человек из 10 тыс. является альбиносом, при этом признак альбинизма у человека определяется одним рецессивным геном. Давайте вычислим, какова доля скрытых носителей этого признака в человеческой популяции. Если один человек из 10 тыс. является альбиносом, то это значит, что частота рецессивных гомозигот составляет 0, 0001, т. е. q2 = 0, 0001. Зная это, можно определить частоту аллеля альбинизма q, частоту доминантного аллеля нормальной пигментации р и частоту гетерозиготного генотипа (2pq). Люди с таким генотипом как раз и будут скрытыми носителями альбинизма, несмотря на то что фенотипически этот ген не будет у них проявляться и они будут иметь нормальную пигментацию кожи.
Из приведенных простых расчетов видно, что, хотя число альбиносов крайне невелико — всего лишь один человек на 10 тыс., ген альбинизма несет значительное количество людей — около 2%. Иными словами, даже если признак фенотипически проявляется очень редко, то в популяции присутствует значительное количество носителей этого признака, т. е. особей, имеющих этот ген в гетерозиготе.
Благодаря открытию закона Харди—Вайнберга процесс микроэволюции стал доступен непосредственному изучению: о его ходе можно судить по изменениям из поколения в поколение частот генов (или генотипов). Таким образом, несмотря на то что этот закон действителен для идеальной популяции, которой нет и не может быть в природе, он имеет огромное практическое значение, так как дает возможность рассчитать частоты генов, изменяющиеся под влиянием различных факторов микроэволюции. История изучения нуклеиновых кислот В 1847 из экстракта мышц быка было выделено вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5'-фосфатом, и содержит N-гликозидную связь. В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало выраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3. Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации. В 1889 г Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей. Левин и Жакоб, изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили адекватные структурные формулы, описывающие их свойства. В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК»[3], оказавшуюся впоследствии ошибочной. В 1935 году Клейн и Танхаузер с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида. Это открыло новые возможности для установления структуры этих соединений. В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов. В результате их работы были установлены все детали химического строения и стереохимии нуклеотидов. За цикл работ в этой области Александер Тодд был награждён Нобелевской премией в области химии в 1957 году. Чаргаффом было установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа. В 1953 году Уотсоном и Криком установлена вторичная структура ДНК, двойная спираль. Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 1229; Нарушение авторского права страницы