
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Хим. Состав и структура ядра и рибосом. ⇐ ПредыдущаяСтр 9 из 9
Ядро растительной клетки (10 – 30 мкм в диаметре) окружено оболочкой, представляющей двойную мембрану. В ядерной оболочке имеются поры размером 10- 20 нм, связывающие ядро с цитоплазмой. Количество и диаметр пор непосредственны: они зависят от функциональной активности ядра. В ядерном соке присутствует хроматин – вещество, состоящие из белков, ДНК, РНК. В нем сосредоточена наследственная информация. Функции ядра заключаются как в хранении и передаче наследственной информации, так и в регуляции всей жизнедеятельности клетки. Это достигается главным образом путем последовательной экспрессии и репрессии генов в процессе реализации наследственной программы. Ядерная оболочка в определенных местах дает начало мембранам эндоплазматического ретикулума. Ограниченные мембранами каналы пронизывают всю цитоплазму, иногда расширяясь в небольшие вакуоли. ЭПР бывает гладким и шероховатым. Шероховатость ЭПР зависит от того, что на внешней стороне мембраны сосредоточены рибосомы. На гладкой ЭПР рибосом нет. Важнейшую функцию в клетке выполняют рибосомы: в них происходит заключительный этап биосинтеза белков - трансляция. Рибосомы находятся в ядре, а затем через поры ядерной оболочки выходят в цитоплазму. Они состоят из белка и рибосомальной РНК. Рибосомы – очень маленькие частицы, образованные из двух субъединиц. Рибосомы могут находиться непосредственно в гиалоплазме или ассоциироваться с мембраной ЭПР. В синтезе одного белка, как правило, принимают участие несколько рибосом, объединенные одной молекулой информационной РНК. Это способствует более высокой активности белкового синтеза. 86.Методы изучения фотосинтеза. Основные показатели….. Фотосинтез – это сложный физиологический процесс усвоения солнечной энергии растениями и трансформацию в химическую энергию органических соединений, сопровождается выделением О2. Формирование урожая с/ х культур – сложный процесс, результат которого определяется взаимодействием растений с условиями внешней среды. Основными показателями, характеризующими фотосинтетическую деятельность посева являются: КПД ФАР, Листовая площадь, ИЛП, ФСП, ЧПФ, СРП. Максимальный фотосинтез обеспечивается при оптимальной листовой поверхности, начинающееся затенение, листья нижних ярусов находятся в недостаточной освещенности, имеет низкую интенсивность фотосинтеза. Для большинства полевых культур оптимальная площадь листьев 40-50 тыс м2 на 1 га. Для кормовых культур оптимальная площадь выше 60-70 тыс м2 на 1 га. ИЛП (индекс листовой поверхности) – характеризует, во сколько раз площадь листьев больше, чем площадь посева. ФСП (фотосинтетический потенциал посева) – тесно хорервирует с урожайностью, характеризует как размеры листовой поверхности, так и длительность работы. Величина разная для разных культур, сортов. Важно учитывать динамику формирования площади листьев. Чтобы большая часть фотосинтетического потенциала приходилась на период интенсивности фотосинтетического аппарата. Правильно учитывать величину ФСП не только по площади листьев, а и с учетом других зеленых частей растений. ЧПФ – показывает накопление биомассы единицы площади листьев за единицу времени. СРП – скорость роста посева, находится произведением ЧПФ на ИЛП. Механизмы поступления воды в растительную клетку. На долю воды в клетке приходится от 60-80 до 95 % массы. Содержание изменяется в зависимости от типов ткани и ее физиологического состояния. Чем активней представлен метаболизм, тем активней физиол. Состояние. Значение воды определяется ее свойствами: способность быть растворителем; полярность; способность испаряться при любой температуре; удельная теплоемкость; высокое поверхностное натяжение; высокая активность в хим. реакциях. Значение воды: она структкрообзователь; определяет транспорт органических и минеральных веществ; это среда для протекания реакций; участник физиологических процессов; участвует в регуляции температурного режима; влияет на активность ферментов. На процессы синтеза используется примерно 1, 5 % воды проходящей через растения. Водообмен складывается из процессов: поглощения воды; передвижение и распределение; испарение. О состоянии воды в клетке - это свободная, осмотически – поглощенная вода, составляющая вода, активный участник физиологических процессов. Ее содержание определяется устойчивостью растений к неблагоприятным условиям среды. Основными механизмами поглощения воды в клетку являются осмос и набухание. Явление набухания обусловлено колоэтальными и каппилярными эффектами. Этот механизм характерен для поглощения воды семенами и меристемами. Это передвижение диполей воды, вызываемое электрическим потенциалом на мембране. При поглощении воды тратится энергия. Поглощение транспорта осуществляется строго в направлении увеличения водного потенциала. Водный потенциал характеризует реальную возможность поглощать воду – это мера активности воды. Клеточные основы роста. В основе роста растений лежит увеличение числа и размеров клеток, сопровождается их дифференцировкой. Для растений характерен рост клеток специализированный тканью – меристема. Различают опекальную меристему ( апексы стебля и корня); лотеральную ( рост в ширину); вставочную (рост междоузлий). В жизни клетки выделяют 4 фазы: 1) эмбриональная или фаза деления клетки. В этой фазе выделяют собственное деление ( митоз) и периоды между делениями ( интерфаза). В интерфазе происходит репликация ДНК ( самоудвоение), активный рост протопласта и ядра, при интенсивном синтезе белков. 2)фаза растяжения. Ядро находится в относительном покое, быстрое увеличение объема клеток ( увеличение в 50-100 раз). Это связано с формированием вакуоли; увеличивается, растягивает клетку, увеличивается площадь клеточной оболочки за счет активного синтеза целлюлозы и др. компонентов Кл. стенки. На этом этапе важно роль фауксинов. 3) Фазадифференциации. Появляются характерные особенности конкретной специальной ткани. В основе дифференциации клеток находится дифференциальная активность генов. Генная регуляция определяет набор ферментов, который определяет путь развития клетки. 4) Фаза старения и отмирания. Резкое снижение интенсивности всех физиологических процессов. Преобладание процессов гидролиза над синтезом. Причины: 1) включение генетической подпрограммы старения; 2) накопление повреждений в генетическом аппарате в мембранах и других органах и увеличение ядовитых веществ в клетке. 89. Возрастные изменения морфологических свойств …….. В индивидуальном развитии высших растений (онтогенезе) выделяют 5 этапов: эмбриональный; ювенильный или молодости; зрелости; размножения – плодоношения и старости. Каждый этап имеет специфические морфологические признаки, физиологические и биохимические свойства, которые тесно связаны друг с другом и находятся в постоянном взаимодействии. Основным критерием начала очередных этапов принимают возникновение характерных зачаточных структур. Эмбриональный этап – у семенных – это период от оплодотворения яйцеклетки до начало прорастания зародыша; у вегетативно – размножающихся – это процесс формирования клубней, луковиц, корневищ до прорастания почек. В течении этого этапа происходит формирование и развитие зародыша, накопление запасных, питательных веществ и переход семян в состояние покоя. От успешности происхождения этого этапа зависит формирование будущего урожая Формирование зародыша называется эмбриогмнез. 3 фазы в эмбриогенезе злаковых: 1) налив зерна – в основном формируется эндосперм: 2) молочная спелость – преимущественно развивается зародыш; 3) восковая и полная спелость – интенсивное отражение зап. Веществ, быстрое снижение содержания воды, накопление ингибиторов роста. В покоящихся семенах, клубнях происходят физиологические, биохимические процессы, способствующие послеуборочному созреванию. Ювенильный – начинается с прорастания семян или почек органа вегетативного размножения. В прорастании семян выделяются фазы: набухания; проклевывание; гетеротрофный рост проростка; переход к автотрофному питанию. Этап молодости длится до закладки репродуктивных органов. Высокая освещенность или длинный день в начале роста сокращают ювенильный период многих растений. Этап зрелости – начинается с закладки цветочных зачатков, формирования клубней, луковиц и других органов вегетативного размножения, гаметогенез, спорогенез. Этап размножения – начинается с оплодотворения, завязавшиеся семена становятся мощными, атрагирующими центрами. Из прилегающих к нимнематолитов, синтезируются запасные пит. Вещества. Этап старения – период закономерного ослабления процессов жизнедеятельности. На уровне целого растения рост продолжается до глубокой старости, но объем замедляется. Продолжительность жизни растений определяется исходным, генетически обусловленным потенциалом жизнеспособности вида. Для однолетних злаковых растений отмирание целиком, у многолетних трав – ежегодно отмирают генеративные побеги, а узел кущения, корневая система остаются жизнеспособными. У плодовых культур ежегодно после плодоношения отмирают плодовые веточки, осенью опадают листья, но большая часть побегов и корневая система сохраняют жизнеспособность. Причины старения: запрограммированный процесс; результат накопления повреждений в онтогенезе клеток, органов, растений. Онтогенез (жизненный цикл) или индивидуальное развитие, — комплекс последовательных и необратимых изменений жизнедеятельности и структуры растений от возникновения из оплодотворенной яйцеклетки, зачаточной или вегетативной почки до естественной смерти. Ускоряют старость – это недостаток воды и низкая, или высокая температура. Снижают рост – раннее удаление генеративных органов, и оптимальные условия роста. Качество урожая: вегетативный период, период закладки вегетативных органов, формирование семени и его налива, период созревания и влияние внешних факторов. Alex 1В.урожай. масличные – это р семена и плоды которых содержат от 20 до 60% жиров, и являются основным сырьём для получения растительного масла. В семенах сразу после цветения отмечается высокое содержание белков и углеводов, позднее синтезе белков ослабевает и начинается интенсивное превращение углеводов в жиры, он продолжается до полного созревания семян. В первые периоды созревания в маслах содержится много свободных жирных насыщенных кислот, затем число их снижается и увеличивается количество не насыщенных жирных кислот. 2В урожай. В начале плоды мало отличаются по хим составу от листьев и побегов, изменения наступают в начале окрашивания плодов. Содержание сахара увеличивается, далее идёт синтез сахарозы. По мере созревания плодов содержание всех форм N уменьшается в 2 раза, а количество белкового азота снижается в меньшей степени. Хим состав зависит от района выращивания. Плоды выращенные на юге содержат много сахаров, клетчатки, меньше орг кислот. Меньшее количество осадков и высокие t способствуют повышению содержания сахаров. Р и К уд повышают сахаристость и уменьшают орг кислоты. 3В урожай. При созревание идёт отток пит вещ из вегетативных органов к репродуктивным, и их качественная трансформация. Сначала погибают листья, стебли и корни. Пит вещ в семени имеют свойство сохранять сои качества долгое время, а при благоприятных условиях преобразуются и снабжают зародыш пит вещ на первом этапе роста – прорастание. 4В урожай. Образование клубней начинается через месяц после появления всходов. Синтез крахмала начинается с образования клубней. В период отмирания листьев происходит интенсивное клубнеобразование, в это время сахара поступающие в клубни быстро превращаются в крахмал. В конце вег периода приток углеводов снижается, а расход углеводов на дыхание продолжается = содержание крахмала снижается. На юге и и при внесение К и в засуху крахмала больше. 5В урожай. Устойчивые сорта к вредителям болезням дают больший урожай, различные сорта по разному используют уд и солнечную радиацию, т.к. имеют большей фотосинтетический индекс. На плодородной почве урожайность возрастает за счёт более лёгкого доступа к элементам пит. Сроки уборки должны быть оптимальными для сорта, чтобы не было потерь. Близняшки 6.Ионный транспорт в растении: внутриклеточный, ближний, дальний транспорт. В зависимости от уровня организации процесса различают три типа транспорта веществ в растении: внутриклеточный, ближний ( внутри органа ) и дальний ( между органами ). Внутриклеточный транспорт. Передвижение веществ внутри одной клетки осуществляется в результате совместного действия циклозина ( круговое движение цитоплазмы ) и направленной поперек этого движения диффузии, чем может достигаться почти полное перемещение веществ в гиалоплазме. У высших растений движение цитоплазмы происходит при участии сократительных белков актомиозинового типа. Скорость движения цитоплазмы 0.2-0.6 мм/мин. Во внутриклеточном транспорте веществ принимают участие также каналы эндоплазматического ретикулума и везикулы Гольджи. Ближний транспорт. Это передвижение ионов, метаболитов и воды между клетками и тканями внутри органа. Ближний транспорт включает радиальный транспорт веществ в корнях и стеблях, передвижение веществ в мезофилле листьев на небольшие расстояния, измеряемые миллиметрами. Осуществляется он через клетки неспециализированных для транспорта веществ тканей по апопласту – совокупности межклетников и межфибриллярных полостей клеточных стенок, соединительных плазмодесмами и вакууму – дискретной системе вакуолей клеток. Дальний транспорт Это передвижение веществ межу органами растения. Осуществляется по специализированной проводящей системе, вкючающей сосуды и трахеиды ксилемы ( восходящий ток) и ситовидные трубки флоэмы( нисходящий ток ). 7. Физиологическая роль фосфора и серы, их усвояемые формы, поглощение и распределение в растении, внешние признаки недостатка этих элементов в растении. Фосфор. 0, 2-1, 2 % сухой массы. Формы поступления в виде ионов ортофосфорной кислоты 8.Калий, кальций и магний, их роль, усвояемые формы, поглощение и распределение в растении. Внешние признаки недос татка этих элементов. Калий. (0.5-1.2)% сухой массы. Поступает в форме 9.Механизмы поглощения в-в растительной клеткой. Пассивный и активный транспорт вещ-в. Корневая система растений поглощает из почвы как воду, так и питательные ве-ва. Оба эти процесса взаимосвязаны, но осуществляются на основе разных механизмов. Растврор питательных солей не проступает в растение в неизменной концентрации. Из очень разбавленных растворов соли поглощаются быстрее, чем вода, и раствор становится еще более слабым. Наоборот, из концентрированных растворов растение берет больше воды, чем солей, и раствор становится еще более концентрированным.Растение поглощает вещества избирательно, в результате соотношение поглощенных веществ обычно оказывается иным, чем в питательном растворе. Транспорт элементов питания в клетку обеспечивается двумя автономными механизмами – пассивным током ве-в по электрохимическому градиенту и их активным переносом против электрохимического градиента. Так как ионы несут электрохимический заряд, то их распределение между клеткой и средой определяется как разностью электрических потенциалов, так и разностью концентраций. Суммарно эти две величины принято обозначать как электрохимический градиент. Соотношение различных механизмов транспорта элементов питания может изменяться в онтогенезе растений и зависит от многих условий. Так, например, роль пассивного поступления ионов увеличивается при повышении внешней концентрации солей( что возможно в условиях засоления или при локальном внесении удоб-й )Передвижение ионов по градиенту электрохимических потенциалов, определяемых зарядом ионов и разностью концентраций, пассивно, а против него- активно. 10.Ассимиляция нитратного азота. Причины накопления высоких уровней нитратов в раст-х и пути снижения их содержания. Азот входит в состав органических соединений только в восстановленной форме. Поэтому включение нитратов в обмен веществ начинается с их восстановления, которое может осуществляться и в корнях, и в листьях. Относительная доля участия этих органов в первичной ассимиляции нитратов являются видовым признаком. В связи с этим выделяют три основные группы растений. 1. Растения, практически полностью восстанавливающие нитраты в корнях и транспортирующие азот к листьям в органической форме. К этой группе относится горох, люпин, черника, многие древесные растения. 2. Растения практически не прявляющие нитратредуктазной активности в корнях и ассимилирующие подаваемые с пасокой нитраты в листьях. Это дурнишник, сахарная свекла, хлопчатник. 3. Растения, способные восстанавливать нитраты как в корнях, так и в листьях. Это наиболее многочисленная группа, включающая хлебные злаки, кукурузу, фасоль, овощные культуры. У них как правило, восстановление нитратов активнее протекает в листьях, однако доля участия разных органов сильно варьирует в зависимости от обеспеченности растений нитратами, концентрации в среде и интенсивности поглощения ионов аммония и калия, уровня освещенности, температуры и других факторов. Восстановление нитрата осуществляется в два этапа. Восстановление нитрата да нитрита, сопряженное с переносом двух электронов и катализируемое ферментом нитратредуктазой (НР): NO3 НР/2 е NO2 ( в цитоплазме ). Восстановление нитрита до аммиака путем переноса шести электронов и катализируемое ферментом нитратредуктазой ( Ни Р): NO2 НиР/6е NH4( в пластидах ). Восстановление нитратов до нитритов происходит в цитозоле клеток корня и листа. Не исключается возможность локализации нитратредуктазы на плазмалемме и на мембранах органелл, граничащих с растворимой фазой клетки. Но эти ассоциации фермента с мембранами удерживаются слабыми связями и легко разрушаются при выделении органелл. В листьях нитратредуктаза локализована в хлоропластах и в качестве донора электронов использует восстановленный в световой фазе фотосинтеза ферродоксин. В корнях No2 восстанавливается в пропластидах с использованием НАДФ *Н, образовавшегося в пентозофосфатном пути дыхания. Использование разных источников приводит к существенной разнице в энергозатратах на превращение NO3 – в NH4+ в надземных и подземных органах. 12. Физиологическая роль микроэлементов. М/Э относятся к группе незаменимых питательных элементов, содержание которых в растительных тканях измеряется тысячными и стотысячными долями процента. Несмотря на то что м/э требуются в очень малых количествах, в их отсутствие нормальная жизнедеятельность становится невозможной. Недостаток м/э вызывает серьезные физиологические расстройства и нередко приводит к гибели растений уже в раннем возрасте. Объясняется это тем что м/э главным образом функционируют в регуляторных системах клетки. Они выступают в качестве простетических групп ферментов или кофакторов –активаторов ферментов. Примеры М\Э: Медь Марганец, Цинк, Молибден, Бор,
Рост и Развитие. 1.Ростовые движения( тропизмы, настии), их природа и значение в жизни растений. О сновой роста тканей, органов и всего раст. явл. образ-е и рост меристематической ткани. Различают апикальную, лотеральную и интеркалярную(вставочную)меристемы. Апикальная-верхушечная образовательная ткань стеблей и корней. Лотеральная-расп. паралельно боковой поверх-и органа. Интеркалярная- в междоузлиях стебля и в основании листьев раст. злаков. Клетка проходит ряд последоват-ых этапов(фаз)своего роста и развития: деления(эмбриональная), роста растяжением(растяжения), дифференцировки (дифференциация), старения и смерти. Движение раст. изм-е положения органов раст. в прост-ве, обусловленное разн. факторами вн. среды: светом, t, силой тяж., хим.элементами и др. Тропизмы-ростовые движения, вызываемые односторонним действием раздражителем. Изгибание раст. в сторону действующего фактора наз.положит. тропизм, а в пртивоп-ю от дейст. фактора-отрицат. В зависимости от природы раздражителя, вызывающего изгиб. каждый конкретный тропизм получил название. Если изгиб вызван действием света- фототропизм, силой тяжести- геотропизм, неравномерным распр. Воды в почве- гидротропизм, хим. соед.(удобрений)- хемотропизм, кислорода-аэротропизм. Настии- изменения положения органов прикрепленных раст., вызванные внешними раздражителями. Настии проявляются благодаря изменениям тургорного давления. Названия, как и тропизмов, зависят от тех раздражителях, кот. их вызывают(фото-, термо-, хемо-, гидро-, тигмо-, сейсмо-, электро- и трвмонастии.) ДЫХАНИЕ РАСТЕНИЙ 7. Дыхание – обеспечивает энергией все процессы жизнедеятельности: - поддерживает структуру цитоплазмы -дыхание роста -поглощение -транспорт -выделение -мех. работа - источник промежуточных вещ-в для различных биосинтезов -источник энлогенной воды Митохондрии ограничены двумя мембранами. Внешняя мембрана хорошо проницаема для неорганических ионов и относительно крупных молекул орг. соед. внутренняя мембрана ограничивает внутреннее содержимое митохондрии, ее матрикс, состоящий на половину из белка. Именно здесь локализованы ферменты цикла Кребса, ферменты, катализирующие окисление жирных кислот, а так же РНК и ДНК для биосинтеза белка. Характерной чертой внутренней мембраны являются складки, кот. Имеют вид плоских гребней, или крист, увеличивающих внутреннюю поверхность митохондрий. Основная ф-ция митохондрий – обеспечение энергетических потребностей клетки. 6. Дегидрогеназы. Основная ф-ция – активирование водорода в молекуле дыхательного субстрата и отделение его от окисляемого вещ-ва, или дегидрирование. Анаэробные дегидрогеназы – передают активированный водород непосредственно на кислород.Этодвухкомпонентные ферменты, коферментом которых является НАД или НАДФ. Аэробные дегидрогеназы – передают активированный водород на промежуточный переносчик. Это двухкомпонентные ферменты, куда наряду с белком в качестве простетической группы входит рибофлавин – производное витамина В2. Оксидазы – Они активируют кислород и катализируют его присоединение к различным органическим соединениям (аминокислоты, фенолы, ненасыщенные жирные кислоты, ксенобиотики – чужеродные токсичные вещества). 4. Анаэробная фаза дыхания – гликолиз проходит в гиалоплазме, где предварительно активированная глюкоза расщепляется и частично окисляется до пировиноградной кислоты ((ПВК). На активирование глюкозы затрачивается энергия 2 молекул АТФ. Активированная глюкоза распадается на 2 молекулы фосфоглицеринового альдегида (ФГА), кот. Окисляются до фосфоглицериновой кислоты (ФГК). Акцептором водорода и электронов в этой реакции служит НАД. Окисление сопровождается перераспределением внутримолекулярной энергии, возникновением макроэнергитической связи, и переносом ее на АДФ с образованием АТФ. Вторым пунктом образования АТФ является р-ция дегидротации фосфоглицериновой кислоты до фосфоленолпировиноградной кислоты (ФЕП). Таким образом, энергетический выход анаэробной стадии дыхания составляют 2НАДФ и АТФ. Кроме того, здесь образуются промежуточные продукты, необходимые для биосинтеза глицерина – компонента жиров, лигнина, фенольных соед-й. 5. Процесс фосфорилирования АДФ с образованием АТФ, сопряженный с транспортом электронов от окисляемого субстрата к О2, получил название окислительного фосфорилирования. Этот процесс является основным источником энергии АТФ в клетке, требующим достаточно сложной структуры как ЭТЦ, так и самих митохондрий. Реакции, в которых энергия, освобождающаяся на определенных окислительных этапах брожения запасается в молекулах АТФ, получили название субстратного фосфорилирования. Их особенностью является катализирование растворимыми ферментами. Образующийся в восстановительной части окислительно-восстановительных преобразований сбраживаемого субстрата восстановитель переносит электроны на подходящий эндогенный акцептор электрона или освобождается в виде газообразного водорода. 8. внутренние факторы: - субстраты дыхания и дыхательный коэффициент. Субстратом для дыхания растений могут служить не только глюкоза, но также жиры и белки, кот. предварительно должны быть подвергнуты гидролизу. Для характеристики дыхательного субстрата используется коэффициент – отношение объема выделившегося СО2 к объему поглощенного О2. - содержание углеводов -донорно-акцепторные отношения. Характер регуляции дыхания определяется соотношением роста и фотосинтеза растении или связями между акцепторными и донорными тканями в конкретных условиях выращивания. Потребности акцептора (роста) являются при этом доминирующими. -возраст растения и его органов. Более молодые растущие органы и ткани дышат интенсивнее. Интенсивность дыхания листьев и корней максимальная в молодом возрасте и снижается по мере уменьшения скорости роста. Внешние факторы: - температура. Почки лиственных пород и иглы хвойных могут дышать при 20-25С мороза. Интенсивность дыхания быстро возрастает при повышении температуры до 35-40С, дальнейшее увеличение температуры приводит к снижению дыхания. - кислород. 21процент в атмосфере – достаточно для окисления. Минимальное сод. кислорода в почве 5процентов, 10-12 – оптимально - вода не только среда, но и участник р-ций - мин.питание - концентрация СО2 и О2. Удвоенное содержание СО2 в воздухе может увеличить урожайность с/х культур на 30-40 процентов. При хранении семян и сочных плодов, если не соблюдается основные условия хранения может значительно уменьшиться сухая масса их вследствие усиленного дыхания. Сохранность зерна регулируется его влажностью. При влажности ниже критической (12-14 процентов) в семенах нет свободной воды и дыхание сведено к минимуму. Главным условием хранения зерна является его влажность, кот. не должна превышать 12процентов от сухой массы. При большей влажности усиливается дыхание, и зерно теряет часть запасных веществ. Минеральное питание. 1) Поглощение элементов минерального питания растением. Растения поглощают вещества избирательно, в результате соотношение поглощенных веществ обычно оказывается иным, чем в питательном растворе. Поглощение та же идет против градиента концентраций. Два этапа поглощения: 1) поступление ионов в свободное пространство корня, 2) транспорт их в протопласты клеток. Первоначальное быстрое поглощение веществ осуществляется в клеточных стенках и является адсорбцией, а быстро выделение – десорбцией. Часть объема корня, где происходят процессы обменной адсорбции – кажущееся свободное пространство (КСП). В почве корень контактирует с почвенным раствором и с частицами почвы, которые являются катионообменниками (ППК). Ионы и различные соединения могут преодолевать мембрану по электрохимическому градиенту (простая диффузия) или через гидрофильные поры, а так же за счет облегченной диффузии гидрофильных веществ. Мембранное строение обеспечивает избирательность поглощения. Наиболее активно поглощают вещества клетки зоны растяжения и корневых волосков.
2) Особенности питания растений аммонийными и нитратными солями. Один из важных факторов поглощения растениями неорганических форм азота – реакция питательной среды. При pH 5 лучше поглощаются нитраты, при pH 7 – аммоний. Аммоний сразу после поглощения метаболизируется в корнях, превращаясь в азот аминокислот и амидов. Группы по способности ассимилировать аммиачный раствор (основа классификации – соотношение между углеводами и белками в семени): наиболее успешно питание идет у растений, у которых углеводы значительно преобладают над белками. Поступившие нитраты запасаются в вакуолях клеток корня, либо подаются с патокой в надземную систему. Метаболизация нитратов начинается с их восстановления до аммония. Аммонийное питание выгоднее нитратного. На поглощение нитратов и аммония сильно влияет температура среды: при пониженной температуре больше поглощается аммония, чем нитрата. Предпочтительность использования форм азота зависит от pH, ионного состава среды, температуры и др. 3) Физиологическая роль азота. Особенности азотного питания растений. Пути снижения и накопления нитратов в составе растительной продукции. Высшие растения не способны усваивать молекулярный азот атмосферы. Это делают азотфиксирующие микроорганизмы. Свободноживущие азотфиксаторы почв бактерии родов Azotobacter и Beijerinckia. Симбиотические азотфиксаторы, поселяющиеся на корнях бобовых растений, вносят больший вклад. Симбиоз – взаимоотношения между высшими растениями и клубеньковыми бактериями. Благодаря клубеньковым бактериям бобовые не только обеспечиваются азотсодержащими соединениями, но и обогащают почву азотом за счет корневых выделений. В основном органический азот почвы усваивается после его минерализации. Основными усвояемыми формами азота для высших растений являются ионы аммония и нитрата. Азот – единственный элемент, который корни растений могут поглощать в форме ионов обоих знаков заряда: анион нитрата NO3 и катион аммония NH4. 4) Распределение по органам, накопление и вторичное использование (реутилизация) элементов минерального питания у растений. Физиологические основы диагностики минерального питания. Направление движения минеральных веществ определяется интенсивностью их использования и потребностями того или иного органа. Реутилизация – повторное использование элементов минерального питания. Наиболее подвижен калий (роль в регуляции транспортных процессов и биоэлектрических явлений). Хорошо реутилизируются азот и фосфор. По мере старения листьев они направляются к более молодым частям растения. Есть элементы, которые практически не реутилизируются. Это кальций, бор, железо (связано с малой лабильностью и плохой растворимостью соединений). Магний и сера промежуточное положение по реутилизации. Для элементов, подверженных реутилизации, характерен базипетальный градиент распределения, т. е. чем выше расположен лист, чем он моложе, тем больше в нем азота, фосфора и калия. Для элементов, не подвергающихся повторному использованию, характерен акропетальный градиент: чем старше орган, тем больше содержание в нем указанных элементов. Благодаря реутилизации растущие части растения на поздних фазах роста получают значительную часть необходимых им элементов.
5) Физиологические основы применения удобрений. Система удобрения – программа удобрений в севообороте с учетом предшественников, плодородия почвы, климатических условий, биологических особенностей растений, состава удобрений. Внесение в почву удобрений приводит не только к увеличению количества золы и азота, но и к увеличению общего количества органических веществ. Минеральные вещества способствуют использованию продуктов фотосинтеза на новообразования. При улучшении водоснабжения путем орошения и в связи с эти усилении ростовых процессов растения больше реагируют на внесение соответствующих удобрений. Растения имеют периоды максимального потребления питательных веществ (в сжатые сроки поступает большое количество минеральных элементов), критические периоды потребления элементов питания (период прорастания). Потребность в элементах минерального питания также тесно связана с продолжительностью вегетационного периода сельхоз культур. При совместном применении минеральных и органических удобрений, увеличении запасов гумуса в почве можно разрешить противоречия необходимости увеличения производства с/х продукции и сохранения окружающей среды от загрязнения. 14 Закономерности роста развития…. Популярное: |
Последнее изменение этой страницы: 2016-04-11; Просмотров: 646; Нарушение авторского права страницы
 :
:  :
:  :
: 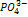 - мало в почве. Находится в нуклеиновых кислотах, фосфолипидах, белеах, нуклеотидов, АТФ, фосфорный эфир сахара, витамины. Вест фосфорный обмен сводится лишь к присоединению или переносу остатка фосфорной кислоты ( фосфорилирование и трансфосфорилирование. Фосфорилирование – это присоединение осттака фосфорной кислоты к какому – либо органическому соединению с образованием эфирной связи. Трансфосфорилирование – процесс при котором остаток фосфорной кислоты, включенный в состав одного орг. в-ва, переносятся на другое ор.в-во. Высока роль в энергетике клетки. В семенах запасная форма-фитин, 6 атомов фосфора в 1 молекуле. Недостаток -активизирует промессы распада орг. соединений, тормозит синтез белков. Тормозится развитие. Характерное изменение окраски листьев с пурпурным или бронзовым оттенком, нижние листья. Легко реутилизируется в форме
- мало в почве. Находится в нуклеиновых кислотах, фосфолипидах, белеах, нуклеотидов, АТФ, фосфорный эфир сахара, витамины. Вест фосфорный обмен сводится лишь к присоединению или переносу остатка фосфорной кислоты ( фосфорилирование и трансфосфорилирование. Фосфорилирование – это присоединение осттака фосфорной кислоты к какому – либо органическому соединению с образованием эфирной связи. Трансфосфорилирование – процесс при котором остаток фосфорной кислоты, включенный в состав одного орг. в-ва, переносятся на другое ор.в-во. Высока роль в энергетике клетки. В семенах запасная форма-фитин, 6 атомов фосфора в 1 молекуле. Недостаток -активизирует промессы распада орг. соединений, тормозит синтез белков. Тормозится развитие. Характерное изменение окраски листьев с пурпурным или бронзовым оттенком, нижние листья. Легко реутилизируется в форме 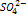 .Нужна особенно бобовым и крестоцветным. В раст. восстанавливается до
.Нужна особенно бобовым и крестоцветным. В раст. восстанавливается до  . В молодых листьях сера в основном находится в составе органических соединений, а в старых накапливается в вакуолях в виде сульфата кальция. Важнейшая функция –поддержание определенного уровня окислительно- восстановительного потенциала в клетке, необходимого для нормального функционирования всех ферментных систем. В составе белков, цистеин, метионин, коферменте-А, витаминах. Участвует и формирует ОВП клетки. Недостаток -тормозит синтез аминокислот и белка, снижает фотосинтез и рост раст., листья бледнеют и желтеют сверху вниз.
. В молодых листьях сера в основном находится в составе органических соединений, а в старых накапливается в вакуолях в виде сульфата кальция. Важнейшая функция –поддержание определенного уровня окислительно- восстановительного потенциала в клетке, необходимого для нормального функционирования всех ферментных систем. В составе белков, цистеин, метионин, коферменте-А, витаминах. Участвует и формирует ОВП клетки. Недостаток -тормозит синтез аминокислот и белка, снижает фотосинтез и рост раст., листья бледнеют и желтеют сверху вниз. .Подвижный элемент, весь в ионном виде, 80% в вакуолях, высокое содержание в активных частях раст. Важный осматический компонент кл сока. Активатор или К-фактор многих ферментов. Участвует в движении устиц, транспорте углеводов, способствует усвоению
.Подвижный элемент, весь в ионном виде, 80% в вакуолях, высокое содержание в активных частях раст. Важный осматический компонент кл сока. Активатор или К-фактор многих ферментов. Участвует в движении устиц, транспорте углеводов, способствует усвоению  .Недостаток- замедляется рост, синтез крахмала, пожелтение листьев в низу вверх. Кальций.. 0.2(0.2-3)% сухой массы. Поступает в форме
.Недостаток- замедляется рост, синтез крахмала, пожелтение листьев в низу вверх. Кальций.. 0.2(0.2-3)% сухой массы. Поступает в форме  .Накапливается в старых органах дыхания, т.к почти не утилизируется. Откладывается в вакуолях в виде нерастворимых солей орг. кислот. Нужен для поддержания структуры хромосом и рибосом. Активатор ферментов. Регулирует вязкость цитоплазмы. Высокое содержание в составе серединной пластинки –соединение пектатов СА. Недостаток- приводит к набуханию пектиновых в-в, это ведет к ослизнению кл. стенки., разрушению клетки. Характерно загнивание: корней, участков стебля, плодов. Магний. 2(0.02-0.3)% сухой массы. Поступает в форме
.Накапливается в старых органах дыхания, т.к почти не утилизируется. Откладывается в вакуолях в виде нерастворимых солей орг. кислот. Нужен для поддержания структуры хромосом и рибосом. Активатор ферментов. Регулирует вязкость цитоплазмы. Высокое содержание в составе серединной пластинки –соединение пектатов СА. Недостаток- приводит к набуханию пектиновых в-в, это ведет к ослизнению кл. стенки., разрушению клетки. Характерно загнивание: корней, участков стебля, плодов. Магний. 2(0.02-0.3)% сухой массы. Поступает в форме  .Много в хлорофилле. Активатор многих ферментов(ДНК и РНК полимераза).Поддерживает структуру рибосом, много в составе фитина. Участвует в азотном обмене. Уселяет синтезвитамина А и С. Недостаток -Мраморная, неравномерная окраска листьев внизу вверх, далее хлороз, некроз верхних листьев. Реутилизируется легче серы.
.Много в хлорофилле. Активатор многих ферментов(ДНК и РНК полимераза).Поддерживает структуру рибосом, много в составе фитина. Участвует в азотном обмене. Уселяет синтезвитамина А и С. Недостаток -Мраморная, неравномерная окраска листьев внизу вверх, далее хлороз, некроз верхних листьев. Реутилизируется легче серы.