|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Часть I. Что вы должны знать о строении и принципе работы мышц.Стр 1 из 7Следующая ⇒
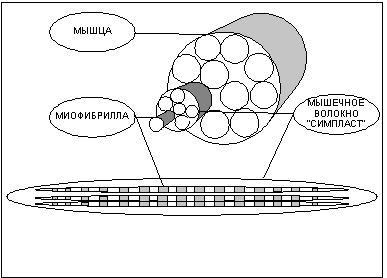
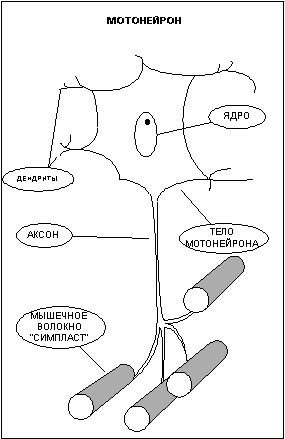
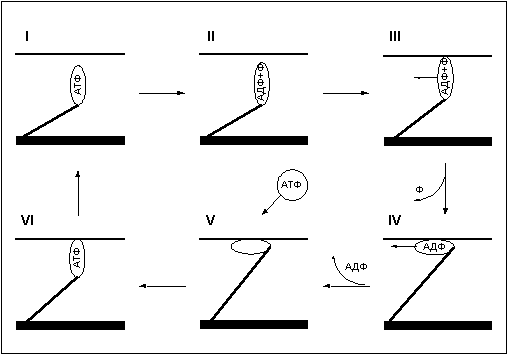
Часть I. Что вы должны знать о строении и принципе работы мышц. Различают три типа мышечной ткани: скелетная, гладкая и сердечная. Функция сердечной ткани понятна из названия, и ее роль, я думаю, объяснять не надо. О существовании гладких мышц мы зачастую даже не догадываемся, так как это мышцы внутренних органов, и мы лишены возможности напрямую управлять ими, впрочем, как и сердечной мышцей. Между тем, именно гладкие мышцы сокращают стенки сосудов, производят сокращение кишечника, способствуя перемещению пищи, и выполняют множество других жизненно важных функций. Задача скелетных мышц - перемещение частей скелета относительно друг друга (отсюда и название). Именно эти мышцы мы с таким упорством пытаемся нарастить на своем теле, и именно их строение и свойства мы будем рассматривать в дальнейшем. Заглянем в клетку. Как известно, все ткани организма имеют клеточную структуру, не представляют исключение и мышцы. Поэтому мне придется провести краткий экскурс в цитологию - науку о клетке, и напомнить читателям о роли и свойствах основных структур клетки. В грубом приближении клетка состоит из двух важнейших, взаимосвязанных между собой частей - цитоплазмы и ядра. Ядро - содержит в себе молекулы ДНК, в которых заключена вся наследственная информация. ДНК - полимер, закрученный в виде двойной спирали, каждая спираль которого составлена из огромного количества четырех видов мономеров, называемых нуклеотидами. Последовательность нуклеотидов в цепочке кодирует все белки организма. Ядро ответственно за размножение клетки - деление. Деление клетки начинается с разделения молекулы ДНК на две спирали, каждая из которых способна достроить парную из набора свободных нуклеотидов и вновь превратится в молекулу ДНК. Таким образом, количество ДНК в ядре удваивается, далее ядро делится на две части, а за ним и вся клетка. Цитоплазма - это все, что в клетке окружает ядро. Она состоит из цитозоли (клеточной жидкости), в которую включены различные органеллы, такие как митохондрии, лизосомы, рибосомы и прочие. Митохондрии - это энергетические станции клетки, в них с помощью различных ферментов происходит окисление углеводов и жирных кислот. Энергия, выделяющаяся при окислении веществ, идет на присоединение третьей фосфатной группы к молекуле Аденезиндифосфата (АДФ) с образованием Аденезинтрифосфата (АТФ) - универсального источника энергии для всех процессов, протекающих в клетке. Отсоединяя третью фосфатную группу и вновь превращаясь в АДФ, АТФ выделяет запасенную ранее энергию. Ферменты или Энзимы - вещества белковой природы в сотни и тысячи раз увеличивающие скорость протекания химических реакций. Практически все жизненно важные химические процессы в организме происходят только в присутствии специфических ферментов. Лизосомы - округлые пузырьки, содержащие около 50 ферментов. Лизосомные ферменты расщепляют поглощенный клеткой материал и собственные внутренние структуры клетки (автолизис). Лизосомы, сливаясь в фагосомы, способны переваривать целые органеллы, подлежащие дезинтеграции. Рибосомы - органеллы, на которых происходит сборка белковой молекулы. Клеточная мембрана - оболочка клетки, она обладает избирательной проницаемостью, то есть способностью пропускать одни вещества и задерживать другие. Задача мембраны сохранять постоянство внутренней среды клетки. Строение мышцы. Структурно-функциональной единицей скелетной мышцы является симпласт или мышечное волокно - огромная клетка, имеющая форму протяженного цилиндра с заостренными краями (в дальнейшем под наименованием симпласт, мышечное волокно, мышечная клетка следует понимать один и тот же объект). Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра. Мышечное волокно, как и любая клетка, окружено оболочкой - сарколемой. Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные и лимфатические сосуды, а так же нервные волокна. Группы мышечных волокон образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в плотный чехол соединительной ткани, переходящей на концах мышцы в сухожилия, крепящиеся к кости. Усилие, вызываемое сокращением длины мышечного волокна, передается через сухожилия костям скелета и приводит их в движение. Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов (рис.2) - нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления - аксоны в составе двигательного нерва подходят к мышце. Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну. Таким образом, один мотонейрон иннервирует целую группу волокон (так называемая нейромоторная единица ), которая работает как единое целое. Рис.2 Мышца состоит из множества нервномоторных единиц и способна работать не всей своей массой, а частями, что позволяет регулировать силу и скорость сокращения. Для понимания механизма сокращения мышцы необходимо рассмотреть внутреннее строение мышечного волокна, которое, как вы уже поняли, сильно отличается от обычной клетки. Начнем с того, что мышечное волокно многоядерно. Связано это с особенностями формирования волокна при развитии плода. Симпласты (мышечные волокна) образуются на этапе эмбрионального развития организма из клеток предшественников - миобластов. Миобласты (неоформленные мышечные клетки) интенсивно делятся, сливаются и образуют мышечные трубочки с центральным расположением ядер. Затем в мышечных трубочках начинается синтез миофибрилл (сократительных структур клетки см. ниже), и завершается формирование волокна миграцией ядер на периферию. Ядра мышечного волокна к этому времени уже теряют способность к делению, и за ними остается только функция генерации информации для синтеза белка. Но не все миобласты идут по пути слияния, часть из них обособляется в виде клеток-сателлитов, располагающихся на поверхности мышечного волокна, а именно в сарколеме, между плазмолемой и базальной мембраной - составными частями сарколемы. Клетки-сателлиты, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни, что обеспечивает увеличение мышечной массы волокон и их обновление. Восстановление мышечных волокон при повреждении мышцы возможно благодаря клеткам-сателлитам. При гибели волокна, скрывающиеся в его оболочке, клетки-сателиты активизируются, делятся и преобразуются в миобласты. Миобласты сливаются друг с другом и образуют новые мышечные волокна, в которых затем начинается сборка миофибрилл. То есть при регенерации полностью повторяются события эмбрионального (внутриутробного) развития мышцы. Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме (в мышечном волокне ее принято называть саркоплазмой) тонких волоконец - миофибрилл (рис.1), расположенных вдоль клетки и уложенных параллельно друг другу. Число миофибрилл в волокне достигает двух тысяч. Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность - чередующиеся темные и светлые полосы. При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой. Энергетика мышцы. Естественно, что для движения мостика требуется энергия. Как я уже упоминал ранее, универсальным источником энергии в живом организме является молекула АТФ. Под действием фермента АТФазы АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (Н3РО4), и превращается в АДФ, при этом высвобождается энергия. АТФ + H2O = АДФ + H3PO4 + энергия. Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения. Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения. Мышца имеет три источника воспроизводства энергии: расщепление креатинфосфата; гликолиз; окисление органических веществ в митохондриях. Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ. АДФ + креатинфосфат = АТФ + креатин. Эта реакция получила название - реакции Ломана. Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников - гликолиза и кислородного окисления. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются. Гликолиз - процесс распада одной молекулы глюкозы (C6H12O6) на две молекулы молочной кислоты (C3H6O3) с выделением энергии, достаточной для " зарядки" двух молекул АТФ, протекает в саркоплазме под воздействием 10 специальных ферментов. C6H12O6 + 2H3PO4 + 2АДФ = 2C3H6O3 + 2АТФ + 2H2O. Гликолиз протекает без потребления кислорода (такие процессы называются анаэробными) и способен быстро восстанавливать запасы АТФ в мышце. Окисление протекает в митохондриях под воздействием специальных ферментов и требует затрат кислорода, а соответственно и времени на его доставку. Такие процессы называются аэробными. Окисление происходит в несколько этапов, сначала идет гликолиз (см. выше), но образовавшиеся в ходе промежуточного этапа этой реакции две молекулы пирувата не преобразуются в молекулы молочной кислоты, а проникают в митохондрии, где окисляются в цикле Кребса до углекислого газа СО2 и воды Н2О и дают энергию для производства еще 36 молекул АТФ. Суммарное уравнение реакции окисления глюкозы выглядит так: C6H12O6 + 6O2 + 38АДФ + 38H3PO4 = 6CO2 + 44H(2)О + 38АТФ. Итого распад глюкозы по аэробному пути дает энергию для восстановления 38 молекул АТФ. То есть окисление в 19 раз эффективнее гликолиза. Типы мышечных волокон. Скелетные мышцы и образующие их мышечные волокна различаются по множеству параметров: скорости сокращения, утомляемости, диаметру, цвету и т.д. Традиционно выделяют красные и белые, медленные и быстрые, гликолитические и окислительные волокна. Скорость сокращения мышечного волокна определяются типом миозина. Изоформа миозина, обеспечивающая высокую скорость сокращения, - быстрый миозин характеризуется высокой активностью АТФазы, а соответственно и скоростью расхода АТФ. Изоформа миозина с меньшей скоростью сокращения - медленный миозин, характеризуется меньшей активностью АТФазы. Волокна, с высокой активностью АТФазы и скоростью расхода АТФ, принято называть быстрыми волокнами, волокна, характеризующиеся низкой активностью АТФазы и меньшей скоростью расхода АТФ, - медленными волокнами. Для восполнения затрат энергией мышечные волокна используют окислительный либо гликолитический путь образования АТФ. Окислительные, или красные, мышечные волокна небольшого диаметра окружены массой капилляров и содержат много белка миоглобина (именно наличие этого белка придает волокнам красный цвет). Многочисленные митохондрии красных волокон имеют высокий уровень активности окислительных ферментов. Мощная сеть капилляров необходима для доставки с кровью большого количества кислорода, а миоглобин используется для транспортировки кислорода внутри волокна от поверхности к митохондриям. Энергию красные волокна получают путем окисления в митохондриях углеводов и жирных кислот. Гликолитические, или белые, мышечные волокна имеют больший диаметр, в их саркоплазме содержится значительное количество гранул гликогена, митохондрии не многочисленны, активность окислительных ферментов значительно уступает активности гликолитических. Гликоген, его еще принято называть " животным крахмалом", - сложный полисахорид с высокой молекулярной массой служит резервным питательным веществом белого волокна. Гликоген распадается до глюкозы, которая, служит топливом при гликолизе. Быстрые волокна, обладающие высокой активностью АТФазы и соответственно скоростью расхода энергии, требуют высокой скорости воспроизводства АТФ, обеспечить которую может только гликолиз, так как, в отличие от окисления, он протекает непосредственно в саркоплазме и не требует времени на доставку кислорода к митохондриям и доставку энергии от них к миофибриллам. Поэтому быстрые волокна предпочитают гликолитический путь воспроизводства АТФ и соответственно относятся к белым волокнам. За высокую скорость получения энергии белые волокна платят быстрой утомляемостью, так как гликолиз, как видно из уравнения реакции ведет к образованию молочной кислоты, накопление которой повышает кислотность среды и вызывает усталость мышцы и в конечном итоге останавливает ее работу. Медленные волокна, характеризующиеся низкой активностью АТФазы, не требуют столь быстрого восполнения запасов АТФ и для обеспечения потребности в энергии используют путь окисления, то есть относятся к красным волокнам. Благодаря этому медленные волокна являются низко утомляемыми и способны поддерживать относительно небольшое, но длительное напряжение. Существует и промежуточный тип волокон с высокой АТФазной активностью, и окислительно-гликолитическим путем воспроизводства АТФ. Тип мышечного волокна зависит от мотонейрона его иннервирующего. Все волокна одного мотонейрона принадлежат к одному типу. Интересный факт - при подключении к быстрому волокну аксона медленного мотонейрона и наоборот, волокно перерождается, меняя свой тип. До недавнего времени существовало две точки зрения на зависимость типа волокна от типа мотонейрона, одни исследователи полагали, что свойства волокна определяются частотой импульсации мотонейрона, другие, что эффект влияния на тип волокна определяется гормоноподобными веществами поступающими из нерва (эти вещества до настоящего времени не выделены). Исследования последних лет показывают, что обе точки зрения имеют право на существование, воздействие мотонейрона, на волокно осуществляется обоими способами. Как строится белок Молекула белка представляет из себя цепочку аминокислот, число звеньев в которой исчисляется от нескольких десятков до нескольких тысяч. Всего в природе насчитывается более 300 видов аминокислот, но для строительства белка используется только 20. Свойства белка определяются последовательностью аминокислот в цепочке, а так же пространственной конфигурацией самой цепочки (вторичная и третичная структура белка). Все белки человека строятся в клетках самостоятельно из аминокислот, поступающих в организм с белковой пищей и синтезируемых самим организмом. Упрощенно процесс синтеза белка изображен на рисунке 7. Код каждого белка записан в ДНК ядра клетки в виде цепочки нуклеотидов. Комбинация из трех нуклеотидов, называемая кодоном, кодирует одну аминокислоту. Последовательность кодонов в ДНК определяет последовательность аминокислот в белке. Ген - последовательность нуклеотидов, кодирующая один белок. Эта последовательность считывается с ДНК и записывается в матричной - РНК (м-РНК), этот процесс называется транскрипцией. мРНК - это как бы кусочек ДНК способный выходить из ядра в саркоплазму, где закрепляется на рибосомах. Транспортные РНК (тРНК) доставляют к мРНК аминокислоты. Один конец тРНК узнает на мРНК соответствующий кодон и прикрепляется к нему. Аминокислота, находящаяся на другом конце тРНК, сцепляется с аминокислотой соседней тРНК, таким образом, выстраивается цепочка белка. Синтез белка очень сложный процесс и его интенсивность зависит от огромного количества факторов. Прежде всего, считывание мРНК в ядре начинается под воздействием стероидных гормонов, вырабатываемых железами внутренней секреции и поступающими в кровь. Молекула гормона из крови проникает в клетку, где, с помощью белка рецептора, доставляется в ядро и разблокирует участок цепочки ДНК, ответственный за тот или иной белок, после чего становится возможной транскрипция мРНК. Для запуска транскрипции РНК, необходимо так же развернуть спираль ДНК, для чего используется фермент РНК-полимераза. На синтез белка сильнейшее влияние оказывает гормон роста (СТГ или соматотропин). По химическому составу соматотропин это белок, поэтому он не может свободно проникать в клетку, в отличие от стероидных гормонов, а воздействует на рецепторы, расположенные на поверхности клетки. Механизм действия гомона роста до конца не изучен, но точно известно, что он стимулирует деятельность РНК-прлимераз и рибосомного аппарата клетки. Ну и конечно для сборки белка требуется наличие в клетке достаточного количества аминокислот и запасов энергии. Без аминокислот не из чего будет строить белок, а энергия нужна для сборки молекулы. И так, для успешного синтеза белка требуются, как минимум, следующие условия: Теперь осталось только ответить на вопрос - как именно тренировка влияет на синтез белка? Должен разочаровать читателя детально объяснить механизм этого влияния на сегодняшнем уровне развития науки невозможно. Если о том, как происходит регуляция синтеза белка в простейших одноклеточных организмах, когда в клетке может идти строительство всех белков, закодированных в ДНК, ученые имеют определенное представление, то, как осуществляется регуляция синтеза белка в многоклеточных организмах, когда, теоретически, каждая клетка может синтезировать все возможные белки, закодированные в ДНК, но синтезирует лишь набор белков, присущий данному типу клеток, остается пока не известным. Да гормоно-рецепторный комплекс разблокирует участок ДНК, в котором закодирован определенный белок, но как гормон узнает какой именно ген в данный момент необходим клетке - миозин быстрого волокна или миозин медленного волокна, а может быть миоглобин? Генетикам предстоит еще долгий путь, прежде чем раскроются все тайны синтеза белка. А как же быть до тех пор? На сегодняшний момент существует несколько гипотез, пытающихся объяснить влияние тренировки на синтез белка в мышце. Но все эти гипотезы можно объединить в два конкурирующих направления - теория накопления и теория разрушения. Суть теории накопления состоит в том, что во время мышечной деятельности в клетке вырабатываются некие факторы-регуляторы, оказывающие влияние на процессы считывания информации с ДНК. Некоторые ученые относят к этим факторам повышение кислотности среды в результате мышечной деятельности, влияющее на спирилизацию ДНК. Некоторые относят к факторам-регуляторам свободный креатин - при интенсивной деятельности кретинфосфат, содержащийся в клетке, в целях восполнения энергии передает свою фосфатную группу на АДФ, превращаясь в креатин, и именно креатин, по мнению ученых, оказывает регулирующее воздействие на ДНК. Думаю, что подобные процессы должны иметь место в регуляции интенсивности белкового обмена, - как известно в случае обездвиживания мышцы интенсивность синтеза белка в клетках снижается, то есть движение само по себе является фактором-регулятором белкового синтеза. Между тем я не могу отвести существенную роль этим процессам в гипертрофии мышц, так как свое регулирующее воздействие данные факторы оказывают непосредственно во время работы мышц, а синтез белка идет восновном после прекращения нагрузки во время отдыха, когда концентрация факторов-регуляторов уже возвращается к уровню, характерному для состояния покоя. Я полагаю, что более полную картину способна сформировать теория разрушения, суть которой заключается в нижеследующем. Как я уже упоминал выше - организм это саморегулируемая система, настроенная миллионами лет эволюции на поддержание постоянства внутренней среды. Разрушение внутренних структур организма автоматически запускает процессы, стремящиеся восстановить утраченное равновесие. Так разрушение белковых структур клетки должно тут активизировать восстановительные процессы синтеза белка, создав все необходимые условия для их протекания. То, что активность синтеза белка в поврежденной ткани в несколько раз выше, чем в нормальных условиях - это факт. Интенсивные восстановительные процессы не могут затихнуть сразу по завершению восстановления поврежденных структур. Как и все прочие процессы, процессы синтеза белка имеют некоторую инерцию, поэтому, в результате восстановления будет наблюдаться некоторый избыточный анаболизм, приводящий к превышению уровня белка в клетке над исходным. Другими словами, будет наблюдаться хорошо известная нам по восстановлению энергетических ресурсов суперкомпенсация. То есть восстановление белковых структур клетки подчиняется тем же общим законам адаптации, с ко торыми вы уже знакомы. Обычно регулирующую роль тренировки в гипертрофии мышц сводят лишь к интенсификации процессов синтеза РНК в ядрах клеток. Между тем общий объем мышцы зависит от количества в ней мышечных клеток/волокон и от количества ядер в мышечных клетках/волокнах (напоминаю, что мышечная клетка и волокно это один и тоже объект). Согласно утвердившимся в среде спортивных физиологов представлении число мышечных клеток/волокон задается генетически и не меняется в ходе тренировок, - об этом свидетельствуют большинство экспериментов, проводившихся в данном направлении (Шекман Б.С.), хотя имеется и ряд экспериментальных данных заставляющих усомнится в этом постулате (об этом чуть позже). Объясняется неизменность количества клеток/волокон в мышце тем, что мышечная клетка представляет из себя сложный многоядерный объект, ядра которого утрачивают способность к делению, как и вся клетка, еще на этапе эмбрионального развития. Между тем потенциальный объем клетки/волокна зависит от количества в ней клеточных ядер - источников РНК. При прочих равных условиях волокно с большим количеством ядер будет иметь больший объем. А теперь внимание! Как показывает ряд экспериментов (M Cabric и N.T.James) в ходе тренировок в мышечных клетках увеличивается количество клеточных ядер. Но ядра мышечных клеток не способны к делению! Так откуда же взялись новые ядра? Ответ на этот вопрос можно найти в работах ученых, занимающихся проблемами регенерации травмированной ткани. Как оказалось, на этапе эмбрионального развития, не все клетки эмбриона, из которых развивается мышечная ткань, сливаются в мышечные волокна и утрачивают способность к делению, часть из них (около 10%) остается в оболочке волокон в виде клеток-сателлитов. Клетки-сателлиты сохраняют способность к делению на протяжении всей жизни и являются резервом восстановления мышечной ткани. Только клетки сателлиты способны быть источником новых ядер в волокне. Как показывают эксперименты (Володина А.В., Женевская Р.П., Климов А.А. и Данилов Р.К., Улумбеков Э.Г. и Челышев Ю.А.) повреждение волокна приводит к активации клеток-сателлитов, которые, освободившись из оболочки, вступают в цикл деления, затем сливаются вместе, восстанавливая поврежденные волокна. Логично предположить, что к активации клеток-сателлитов после тренировки приводят процессы аналогичные травмам волокон. Многие знают на собственном опыте, что интенсивная тренировка, особенно после продолжительного перерыва, отзывается болью в последующие несколько дней отдыха. Боль явно свидетельствует о разрушениях внутренней структуры мышц. Микроскопические исследования показывают, что в результате тренировок в ряде мышечных волокон нарушается упорядоченное расположение миофибрилл, наблюдается распад митохондрий, а в крови повышается уровень лейкоцитов, как при травмах или инфекционном воспалении (Морозов В.И., Штерлинг М.Д с соавторами). Разрушение внутренней структуры мышечного волокна во время тренировки, назовем его микротравмой, приводит к появлению в волокне обрывков белковых молекул, что активизирует лизосомы, " переваривающие", с помощью содержащихся в них ферментов, белковые структуры, подлежащие уничтожению. Если лизосомы не справляются с объемом повреждений, то через сутки наблюдается пик активности боле мощных " чистильщиков" - фагоцитов. Фагоциты - клетки, живущие в межклеточном веществе и крови, основная задача которых уничтожение поврежденных тканей и чужеродных микроорганизмов. Именно продукты жизнедеятельности фагоцитов вызывают воспалительные процессы и боль в мышцах, через сутки после тренировки. Но между тем, по-видимому, именно благодаря деятельности лизосом и фагоцитов повреждается оболочка мышечного волокна, и из нее высвобождаются клетки-сателлиты. Освободившись, клетки-сателлиты начинают цикл деления и сливаются с поврежденным волокном, увеличивая в нем количество ядер, тем самым, повышая его потенциальную возможность в синтезе белка. В свете выше сказанного, я бы не стал полностью исключать возможность высвобождения клеток-сателлит в межклеточное пространство и слияние их в новые волокна, что достоверно наблюдалось в случае обширных повреждений мышечной ткани, правда новые волокна в этих случаях образовывались в замен утраченных, что вовсе не приводило у общему увеличению количества волокон в мышце. Но, если предположить, что повреждения волокна не столь обширны, чтобы привести к его гибели, а клетки сателлиты пошли по пути слияния в новое волокно, то гиперплазия становится не таким уж невероятным фактом, как это принято считать. Тем более что есть ряд экспериментов выбивающихся из общих представлений о невозможности гиперплазии. Так Goneya W, удалось на 19-20% увеличить количество мышечных волокон в лапах кошек, которых он заставлял тренироваться с прогрессирующей нагрузкой. А Yamada S, Buffinger N, Dimario J& Strohman R (1989) и Larson L& Tesch PA (1986) проводили взятия проб из мышечной ткани у элитных бодибилдеров, и контрольной группы людей не обладающих значительной мускулатурой, анализ проб показал, что поперечное сечение волокон у элитных бодибилдеров лишь незначительно больше чем у представителей контрольной группы, в то время как поперечное сечение мышц различалось существенно, то есть бодибилдеры обладают большим количеством волокон по сравнению с контрольной группой, что может быть либо следствием гиперплазии волокон, либо элитные бодибилдеры от рождения обладали значительно большим количеством волокон, чем обычные люди, но эти волокна были крайне тонкими (так как до тренировки элитные бодибилдеры обладали мускулатурой обычных размеров). В последнее хочется верить меньше всего, так как эта теория ставит крест на возможности обычных людей добиться сколько ни будь значительной гипертрофии мышц. Однако не будем зацикливаться на вопросах гиперплазии, и так как возможность последней у человека считается недоказанной, будем, по прежнему, исходить из того, что рост мышц происходит исключительно по причине гипертрофии уже существующих волокон. Но вот одной из причин гипертрофии самих волокон, как раз и является увеличение в них количества клеточных ядер, что по оказываемому эффекту практически равносильно гиперплазии. Вернемся к рассмотрению процессов происходящих в мышце во время восстановления после тренировки. По завершении катаболического этапа саморазрушения поврежденных структур начинается этап компенсации - восстановления внутренней структуры волокон, который, ради справедливости должен заметить, не всегда может завершиться суперкомпенсацией. При слишком обширных травмах или отсутствии условий для восстановления результат может быть прямо противоположным. Против теории разрушения чаще всего приводят следующие аргументы: " Если причиной роста являются микротравмы, то почему же мышца не растет, если ее бить палками? " Ответ на этот вопрос можно найти в работе Володиной А.В., целью ее докторской диссертации является изучение процессов, препятствующих реализации регенерационного потенциала, заложенного в мышечном волокне. Эксперименты показали, что, в условиях обширного повреждения волокон, сопровождающегося ишемией (нарушением кровоснабжения) поврежденных тканей, вызывающей дефицит в снабжении волокна кислородом и питательными веществами, часть клеток-сателлит гибнет и поглощается фагоцитами, а часть идет по пути превращения не в мышечные клетки, а в фибробласты (клетки, производящие коллаген). В итоге место повреждения затягивается соединительной тканью, а количество волокон в мышечной ткани снижается, по причине гибели части из них от повреждений. Очевидно, что при микротравмах волокон - разрушении внутренней структуры волокна без нарушения его целостности, в отличие от травм целой мышцы, снабжение волокон кислородом, а так же его иннервация не нарушены, поэтому условия, приводящие к гибели целых волокон и клеток-сателлитов, отсутствуют. И так, если объем микротравм, полученных в ходе тренировки, был не слишком велик для срыва восстановительных процессов, но достаточен для активации клеток-сателлитов, то в подвергшемся тренировочной нагрузке волокне увеличивается количество клеточных ядер. Восстановление энергетических ресурсов после тренировки приводит к суперкомпенсации энергетических веществ, а лизис разрушенных тренировкой белков увеличивает содержание свободных аминокислот непосредственно в волокне, что в совокупности создает благоприятные условия для интенсификации процессов синтеза белка. При условии достаточного по времени и полноценного отдыха, отсутствия новых стрессовых нагрузок, адекватного снабжения волокна энергией и пластическими ресурсами (аминокислотами) интенсивные процессы восстановления приведут к накоплению в волокне белковых структур сверх уровня, который был до тренировки, то есть будет наблюдаться гипертрофия мышц. Надо отметить, что последовательность протекания фаз общей неспецифической адаптационной реакции (синдрома стресса) такова, что обеспечивает поддержку описанных выше регенерационных процессов на системном уровне. Первая катаболическая фаза стресс реакции, сопровождается выбросом кортикостероидов, что приводит к мобилизации энергетических ресурсов организма и обеспечивает индукцию ферментов лизосом и фагоцитов, расщепляющих белок (кортикостероиды являются теми гормонами, которые активируют на ДНК клеток гены протеолитических ферментов), что способствует скорейшему очищению волокон от поврежденных структур. В последующей фазе стресс реакции синтез кортикостероидов сменяется синтезом анаболических гормонов, что обеспечивает на системном уровне компенсаторный анаболизм. Как тут не вспомнить об основном законе философии - единстве и борьбе противоположностей. Анаболизм активируется катаболизмом - рост мышц есть следствие их предварительного разрушения. Что такое микротравма Открытым остается вопрос, что вызывает разрушение внутренней структуры волокна и является тем самым стрессом для мышцы? Прежде чем ответить на этот вопрос, рекомендую читателям вспомнить механизм сокращения мышц, описанный в I части. Ученый и пауэрлифтер Фредерик Хетфилд, считающий роль микротравм в тренировочном процессе скорее отрицательной из-за необходимости длительного восстановления, полагает, что причиной микротравм является повреждение миофибрильных нитей во время негативных повторений. Вот как он описывает механизм этих повреждений: " Так как количество перекрестных мостиков, старающихся сократить мышцу недостаточно, они буквально " продираются" сквозь мостики соединений нити, стараясь вызвать концентрическое сокращение. Однако сцепиться, как следует им не удается, они срываются и повреждаются. Эти действия, очень напоминающие протаскивание щетины одной зубной щетки через другую, сопровождаются сильным трением, и мышечные нити разрушаются" Не правда ли образно? Но, на мой взгляд, несколько сумбурно - так пишут когда хотят объяснить то, что до конца не понимают сами. К сожалению, мэтр ошибся дважды. Во-первых, микротравмы возникают не только при негативных повторениях, но и при позитивном движении. Во-вторых, использовать термин " трение" для описания взаимодействия молекул некорректно. Сила трения введена в физике для описания на макро-уровне поверхностного взаимодействия тел специально, дабы абстрагироваться от истинной природы " трения" - электромагнитного взаимодействия молекул поверхностного слоя. Механизм повреждения миофибрильных нитей носит несколько иной характер, чем описывает Хетфилд, и мне удалось его аналитически смоделировать. Для понимания механизма повреждения миофибрильных нитей следует обратиться к рассмотрению фаз движения миозинового мостика, которые вкратце уже описаны в первой части статьи. Сейчас остановимся подробнее на этом вопросе (рис. 8). Итак, в первой фазе, еще до сцепления с актином, головка миозинового мостика несет в себе АТФ. Далее во второй фазе под действием фермента АТФаза АТФ гидролизуется, расщепляясь на АДФ и неорганический фосфат. Происходит это на не связанном с актином миозине, после этого миозиновая головка может соединятся с актином - третья фаза. Для совершения рабочего хода мостика используется энергия, освобождающаяся при диссоциации продуктов гидролиза АТФ. Основная доля энергии выделяется при высвобождении неорганического фосфата (переход из третей фазы в четвертую) и меньшая часть при высвобождении АДФ (переход из четвертой фазы в пятую). В пятой фазе -ригорное состояние мостика, мостик уже не генерирует силу, но по-прежнему находится в сцепленном состоянии, вывести его из этого состояния может только молекула АТФ. Поглощая АТФ, головка миозина переходит в шестую фазу, после чего отцепляется от актина, возвращаясь в исходное состояние (первая фаза). Анализируя фазы движения миозинового мостика, я сразу обратил внимание на тот факт, что для отцепления мостика от актина требуется молекула АТФ. При скольжении нитей миозина вдоль актина под действием сил тянущих мостиков (позитивное движение) или под действием внешней силы (негативное движение) сцепленные мостики растягиваются и мешают движению, этим, как вы помните, объясняется различие в силе развиваемой волокном при удлинении и сокращении и сокращении с разной скоростью. Когда АТФ в мышце находится в достаточном количестве, мостики успевают вовремя отцепиться, но что будет, если, при снижении концентрации АТФ в мышце, молекула АТФ не успеет отцепить головку миозина до того, как растяжение мостика превысит предел его прочности? Естественно сцепленный мостик разорвется! (Рис. 9). Популярное:
|
Последнее изменение этой страницы: 2016-08-24; Просмотров: 857; Нарушение авторского права страницы