
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Порядок хлорококковые, или протококковые, СЫогососса1е5-Рго1ососса1е5
Этот порядок объединяет коккоидные формы: одноклеточные, колониальные и ценобиальные. Клетка большинства хлорококковых имеет то же строение, что и у вольвоксовых: в цитоплазме, заполняющей всю полость клетки, располагаются париетальный, часто чщдевидный хроматофор с пиреноидом, одно ядро, отсутствуют жгутики'; стигма и сократительные вакуоли—■ атрибуты монадной организации. У сравнительно немногих представителей хлорококковых (например, у водяной сеточки — гидродикциона) клетка содержит крупную центральную вакуоль, а цитоплазма с хро- матофором и многочисленными ядрами занимает постенное положение. Бесполое размножение —посредством зооспор и апланоспор (автоспор). У ценобиальных форм зооспоры и автоспоры обычно еще внутри оболочки материнской клетки слагаются в дочерние ценобии, которые, как и у вольвоксовых, растут только за счет увеличения размеров клетки, а число клеток остается постоянным.
Половой процесс изо-, гетеро- и оогамный. В зависимости от способа бесполого размножения — зооспорами или - автоспорами — хлорококковые нередко делят на две большие группы: хлорококковые зоо-
Зооспоровые хлорококковые—СН1огососса1ез гоозроппае Род хлорококк (СМогососсит, рис. 81) встречается в пресноводных водоемах, а также на почве, коре деревьев; входит в состав многих лишайников. Представлен шаровидными, микроскопически мелкими клетками, одетыми оболочкой. Клетки одноядерные или многоядериые, содержат чашевидный хроматофор с одним или несколькими пиреноидами. Зооспоры вытянутые, двужгутиковые, образуются в клетке в количестве от восьми до 32 и освобождаются через разрыв в стенке материнской клетки. После некоторого периода движения зооспоры теряют жгутики, одеваются оболочкой и превращаются в новые шаровидные клетки, постепенно дорастающие до размеров материнской (рис. 81). Известен и половой процесс, заключающийся в слиянии дву- жгутиковых изогамет. Род хлорохитриум (СН^о^^сНу^^^ит). Шаровидные, эллипсоидальные или неправильной формы крупные клетки живут в качестве эндо- фитов в межклетниках высших растений (СегШорНуНит, Е1ойеа, Ро- 1атоде1оп, Ритех, ЬусНтз и др.) и внутри таллома морских водорослей (Ро1уз1рНота, РогрНуга, С1дагИпа и др.). Наиболее распространенный вид СН1огосНу1гшт 1етпае (рис. 82) \ I живет в ряске (главным образом Ьетпа 1г1зи1са). Его крупные эллипсоидальные темно-зеленые многоядерные, одетые толстой слоистой оболочкой клетки помещаются в межклетниках ряски. У С1г. 1етпае
Рис. 81. СЫотососсит. А — взрослая клетка; Б— образование зооспор; В — выход зооспор из материнской клетки; Г — зооспора; Д — молодые особи Рис. 82. СШогосНуМшп в ткани ряски
8 Заказ 552 известен только половой процесс1 делений в клетке образуется много показали исследования Л. И. Куреа- нова и его ученицы Н. М. Шемаха- новой, первое из этих делений редукционное. Через разрыв в стенке клетки и окружающей ткани гаметы выходят окруженные широким •слизистым пузырем, внутри которого часто происходит и копуляция, хотя гаметы могут освобождаться из пузыря и до копуляции. Четырехжгу- тиковые зиготы (планозиготы) оседают на новые растения ряски, оде-- ваются оболочкой и образуют труб- ковидный отросток, проникающий
В результате последовательных! 56) двужгутиковых гамет. Как в хозяина или через устьица, или между " эпидермальными клетками. В одном из межклеточных пространств отросток вздувается в эллипсоидальную клетку, которая получает все содержимое планози- готы и медленно растет до наступления новой репродуктивной фазы. Как указывалось, мейоз у СН. 1етпае предшествует гаметогенезу; после слияния гамет восстанавливается диплоидная фаза. Таким образом, СЫогосНу1пит в отличие от остальных хлорококковых и большинства зеленых водорослей является диплонтом: гаплоидны только гаметы. Поскольку хлорохитриум всегда содержит хлорофилл, он питается автотрофно, являясь лишь эндофитом. У близкого к нему рода родохитриум (ЯНос1осНу^^^и^п), распространенного в Америке и обитающего в листьях наземных сложноцветных, эндофитизм развился в паразитизм, так как родохитриум совершенно утратил хлорофилл и питается за счет органических веществ растения, в межклетниках которого он развивается. Род харациум (СНагасшт, рис. 83, А)—одноклеточная эпифит- ная водоросль. Клетки овальные, удлиненные, " веретеновидные, часто асимметричные, прикреплены к субстрату короткой ножкой, расширенной в маленький базальный диск. Размножение двужгутиковыми зооспорами (рис. 83, освобождающимися через разрыв стенки материнской клетки. У некоторых видов наблюдался половой процесс — анизогамия. Клетки превращаются в макро- и микрогаметангии. Первые производят незначительное число крупных зеленых женских гамет, вторые развивают множество мелких желтовато-зеленых мужских гамет (рис. 83, В, Г). Копулируют всегда гаметы, различающиеся по размерам. Род гидродикцион, или водяная сеточка (НуйгосИс1уоп) — макроскопическая, достигающая 30 см и более ценобиальная водоросль. Цилиндрические или- широкоовальные клетки соединяются концами большей частью по три, образуя сеть. У одних видов, например Н. а}г1- сапит, ценобии плоские, несколько загнутые у краев, -1 блюдцевидные (рис. 84, Л); у других, например у широкораспространенной в водах, богатых азотом, Н. геИсиШит, сеть представляет собой замкнутый мешок (рис. 84, Б). Молодые клетки Н. а{псапит цилиндрические, но с возрастом вздуваются, формируя огромные (До 1 см в диаметре) сферы, которые разъединяются и лежат в виде зеленых шариков на дне мелких водоемов, где этот вид встречается. У Н. геИсиШит клетки цилиндрические, с заостренными концами, имеют крупную центральную вакуоль. Постенная цитоплазма содержит многочисленные ядра и сетчатый хроматофор с многочисленными пиреноидами (рис. 84, В). При бесполом размножении в клетке путем последовательного распада протопласта на более мелкие участки возникает много тысяч одноядерных двужгутиковых зооспор. Они движутся некоторое время внутри оболочки материнской клетки, затем втягивают жгутики, выделяют собственную оболочку и соединяются, образуя маленькую дочернюю сеточку (рис. 84, Б), которая освобождается после разрыва стенки материнской клетки.
Как показали новейшие исследования-с применением электронной
микроскопии, зооспоры соединяются не в любом месте их поверхности, а лишь в тех участках, где под плазмалеммой располагаются микротрубочки. Зооспоры, подвергнутые действию колхицина — яда, разрушающего цитоплазматические микротрубочки, не объединяются в ценобий. Молодые сеточки растут только благодаря росту слагающих их клеток, причем число ядер в них увеличивается. Половой процесс изогамный. Двужгутиковые изогаметы образуются так же, как зооспоры, но в большем числе и соответственно они мельче. Гаметы освобождаются через отверстие в стенке материнской клетки. Я. геИсиШит — форма однодомная (гомоталличная: даже гаметы, образовавшиеся в одной клетке, способны копулировать). У гидродикциона изогамия. Однако под электронным микроскопом удалось установить разницу в строении сливающихся гамет: одна из них на переднем конце между жгутиками несет электронно-плотную апикальную шапочку, другая такой шапочки лишена. Апикальная шапочка вытягивается в виде оплодотворяющей трубки, кончик которой и сливается с мембраной, находящейся между жгутиками гаметы без апикальной шапочки (рис. 85). Зигота одевается оболочкой, в ней накапливается жир, окрашенный гематохромом в кирпйчно-красный цвет. Постепенно зигота увеличивается в размерах и переходит в состояние покоя, а затем прорастает, образуя четыре крупные двужгутиковые зооспоры, при этом ядро редукционно делится. Зооспоры, поплавав некоторое время, останавливаются, и каждая развивается в многоугольную клетку — полиэдр. Полиэдр разрастается, становится многоядерным, и содержимое его распадается на двужгутиковые зооспоры, которые слагаются в молодую зародышевую плоскую сеточку, освобождающуюся через разрыв оболочки полиэдра. / Полиэдры, оболочки которых несут отростки, а содержимое богато жировыми включениями, по-видимому, имеют значение для распространения водоросли: крупные зооспоры, освобождающиеся при прорастании зигот, малоподвижны и не могут выполнять эту функцию. Кроме того, полиэдры выносят высыхание, губительное для ценобия, и таким • образом наряду с зиготой могут представлять покоящуюся стадию в жизненном цикле водяной сеточки.
Род педиаструм (РесИаз(гит), имеющий сходный с гидродикционом цикл развития, характеризуется микроскопически мелкими пластинчатыми колониями из разного, кратного двум, числа клеток (от четырех до 128). Клетки, во взрослом состоянии многоядерные, располагаются обычно концентрическими кругами вокруг одной центральной (рис. 86). Краевые клетки часто несут выросты стенки. Двужгутико- вые зооспоры освобождаются через щель в стенке материнской клетки и окружены слизистым пузырем, внутри которого они сначала движутся во всех направлениях. Постепенно они располагаются в одной плоскости и образуют маленькую дочернюю колонию, позднее освобождающуюся. Половой цикл педиаструма такой же, как у гидродикциона, и также включает стадию полиэдра. Автоспоровые хлорококковые — СН1огососса1е$ слйочрог'тае Род хлорелла (СЫогеИа, рис. 87, Л) широко распространен в пресных водах, на сырой земле, коре деревьев и т. д. Шаровидные клетки одеты гладкой оболочкой, содержат обычно чашевидный хроматофор, одно ядро. При бесполом размножении содержимое клетки распадается на четыре или более частей — автоспор, которые еще внутри оболочки материнской клетки одеваются собственными оболочками. Освобождаются автоспоры после разрыва стенки материнской клетки (рис. 87, Б, В). Недавние электронно-микроскопические исследований позволили обнаружить сложное строение клеточной стенки у некоторых видов хлореллы. Наружный трехслойный компонент стенки, образованный двумя электронно-плотными зонами, заключающими электронно-прозрачную центральную зону, содержит спорополленин — чрезвычайно устойчивое к действию различных ферментов вещество, встречающееся в пыльцевых зернах и спорах высших растений. Внутренний, более толстый компонент содержит целлюлозные микрофибриллыПри авто- 1 ряда видов хлореллы в оболочке находится только микрофибриллярный слой, а трехслойный наружный компонент из спорополленина отсутствует. / спорообразовании компоненты стенки развиваются следующим образом. Кнаружи от цитоплазма- тических мембран (плазмалемм) автоспор, заключенных еще в оболочку материнской клетки, появляется первый предшественник клеточной стенки в виде мелких трехслойных изогнутых дисков. Эти диски увеличиваются в числе и размерах, пока краями не сольются, образуя вокруг автоспоры сплошной трехслойный наружный компонент, в котором откладывается спорополленин. Как только сформировался сплошной трехслойный наружный компонент стенки автоспор, начинается растворение внутренней микрофибриллярной части стенки материнской клетки, и между образовавшимся трехслойным чехлом и цитоплазматической мембраной автоспоры откладывается микрофибриллярный компонент стенки автоспор. Стенка материнской клетки, от которой остается 4 только тонкий наружный спо-. г рополлениновый компонент, очень стойкий по отношению к химическим, но не механиче- /кМ(()г§1\ ским воздействиям, разрыва- 1Ш (_______ ) 1Щ ется, и автоспоры освобожда- 1Щ га [Щ ^ Ш1 ются (рис. 88). |П Г) Ш ШккОЮШ/ Другие, близкие к хлорел- ^ш& У ле роды характеризуются иной формой клеток, а также тем, ^^^^^^^ 2 что наряду с одиночно живу- А в щими клетками у них в резуль- тате неполного расхождения ______ автоспор нередко возникают / ^ч^^^Чио скопления клеток или колоний /у щЩь, из неопределенного числа ([ ^тНйй! клеток. Таковы, например, роды анкистродесмус (Лпк1з1го- дезтиз) и кирхнериелла {К1г- сНпепеИа). Первый с верете- новидными, прямыми или изогнутыми клетками (рис. 89, Л), а второй — с полулунными или дуговидными клетками (рис. 89, Б).
. Род эремосфера (Егетозт рНаега), обычный в сфагновых болотах, отличается очень крупными (до 150 мк) одиночными шаровидными клетками с " 'V ^ ■ •■ ЛЬЯ..' Рис, 89. Автоспоровые хлорококковые: А — Апк1& (гойе$пша; Б — К1гсНпег1е11а 1ипапз; В, Г — ЕгетоарНаега штй{$ (В — вегетативная клетка, Г — образование автоспор) одним ядром и многочисленными дисковидными хроматофорами с пиреноидами (рис. 89, В). Автоспоры в числе двух или многих освобождаются через разрыв стенки. Сравнительно недавно, в 1967 г., у Е. ььгШз в экспериментальных условиях наблюдался половой процесс: в одних клетках (оогониях) развивается одна яйцеклетка, в других (спермато- гониях) в результате последовательных делений образуется от 16 до 64 двужгутиковых сперматозоидов. Внутри стенки оогония ооспора одевается скульптурированной оболочкой. Оогамный половой процесс помимо эремосферы гораздо раньше наблюдался у родов голенкиния (Со1епМта), микрактиниум (Мгсга- сНтит) и диктиосфериум (ОШуозркаегшт). Род голенкиния (ОоЫпЫта) представлен свободноживущими шаровидными клетками, стенки которых несут выросты в виде сплошных или полых щетинок (рис. 90, А). У близкого к голенкинии рода микрактиниум (МгсгасИтит) снабженные щетинками шаровидные клетки соединены вуколонии (рис. 90, Г). Не так давно единственным способом размножения голенкинии и микрактиниума, а также всех автоспоровых хлорококковых считалось образование автоспор. Полагали, что автоспоровые хлорококковые утратили в жизненном цикле подвижные жгутиковые стадии, так как бесполое размножение у них осуществлялось неподвижными автоспорами, а половой процесс никогда не наблюдался и считался утраченным. В' 1937 г. русский ботаник А. А. Коршиков впервые наблюдал и описал оогамный половой процесс у новых'описанных им видов Оо1епкШа (О. 1огщ1зр1па, С. зо- Шапа) и у МШ& сМпшт ризШит. В одних клетках протопласт делится с образованием 8—16 двужгутиковых сперматозоидов, в других клетках развивается по одной яйцеклетке. Стенка оогониальной клетки в
нескольких местах ослизняется, и сперматозоид оплодотворяет яйцеклетку. Оплодотворенная яйцеклетка, одетая сначала гладкой оболочкой, выходит из оболочки материнской (оогониальной) клетки и переходит в состояние покоя (рис. 90, Б, В, " Д, Е, Ж). В 1940 г. был описан половой процесс, также оогамный, у другого рода автоспоровых хлорококковых — диктиосфериума (В1с1уо§ркае- гьит). Это колониальная водоросль. Ее клетки (обычно четверками) расположены по периферии слизистых сферических или эллипсоидальных колоний и связаны между собой системой разветвленных нитей, расходящихся из центра колонии и представляющих собой остатки стенок многих генераций клеток. У этого рода при образовании автоспор (обычно четырех) стенка родительской клетки не расплывается, а разрывается на четыре сохраняющиеся лопасти, к концам которых и остаются прикрепленными автоспоры (рис. 90, 3). Достигнув размеров материнской клетки, автоспоры также формируют автоспоры, прикрепленные к лопастям материнской стенки, которые впоследствии скручиваются в нити, и т. д. У одного из видов диктиосфериума, распространенного в Индии (/). иШсит), наблюдался оогамный половой процесс. Этот вид двудомный: можно различать мужские и женские колонии. Клетки мужских колоний производят по 8—16—32 двужгутиковых сперматозоида, которые выходят через поперечный разрыв стенки материнской клетки. Содержимое каждой клетки женской колонии делится только один раз, образуя две округлые яйцеклетки, которые выходят наружу через поперечный разрыв стенки и остаются в слизи, окружающей колонию. Сперматозоиды подплывают к женской колонии и сливаются с яйцеклетками. Ооспора одевается оболочкой и переходит в состояние покоя (рис. 90, И — Н). В деталях оогамия у Э. тсИсит отличается от полового процесса у голенкинии и микрактиниума: у диктиосфериума в каждой клетке образуются по две яйцеклетки (у голенкинии и микрактиниума — по одной) и, кроме того, у диктиосфериума яйцеклетка выходит из оогония до оплодотворения" и сливается со сперматозоидом вне оогония. Такой половой процесс называется примитивной оогамией в отличие от истинной оогамии, при которой яйцо оплодотворяется на месте образования — внутри оогония. Примитивная оогамия очень редко встречается у зеленых водорослей: помимо И. тсНсит примитивная оогамия описана у некоторых вольвоксовых (например, у СЫогоцотит оо§атит, Саг1епа 1уеп§аги) и у одной нитчатой водоросли {СНае1о- пета 1ггеци1аге). Из родов, для которых характерен ценобий, сложенный из определенного числа клеток, различно расположенных, можно рассмотреть С2 и яйцеклетки; Ж—зрелые ооспоры; 3 — Н — ОШуозрНаепит 1п(Исштг. 3 — внешний вид колонии, И — образование в каждой клетке женской колонии по два яйца и'выход их через разрыв клеточной стенки; К—в клетках мужской колонии формируется по 16—32 сперматозоида; Л, М—слияние сперматозоида с яйцеклеткой; Н — зигота Рис. 91. Бсепсйеьтиз диайпсашка. А — ценобий; Б — образование новых ценобиев род сценедесмус (8сепейезтиз), у которого эллипсоидальные или вере- теновидные клетки соединены боками в простые или двойные ряды. У самого обычного вида 5. диайгкаийа (рис. 91, Л), как правило, це- нобии четырехклеточные, причем краевые клетки несут выросты стенки— шипы. При размножении в каждой клетке колонии в большинстве случаев образуются четыре автоспоры, которые внутри оболочки материнской клетки слагаются в молодую колонию (рис. 91, Б). Эта колония позднее освобождается через разрыв материнской стенки и растет только за счет роста клеток, число их не меняется. Электронно-микроскопические исследования многих видов рода 5сепейезтиз показали весьма сложное строение клеточной стенки. Она состоит из трех слоев: внутреннего толстого целлюлозного слоя, очень тонкого среднего слоя, соответствующего трехслойному компоненту стенки СМогеНа и также содержащего спорополленин, и наружного пектинового слоя. Два первых слоя окружают каждую клетку це- нобия, а пектиновый наружный слой одевает весь ценобий, объединяя все его клетки в единое целое. Различная орнаментация клеточных стенок видов сценедесмуса, различимая в оптическом микроскопе и используемая как диагностический признак, обусловлена сложным строением наружного пектинового слоя. Этот слой образован' гомогенным матриксом, на поверхности которого находится сеть, сложенная из шестиугольных ячеек. Сеть покоится на цилиндрических трубчатых под- порках^ основание которых прикрепляется к поверхности среднего (спо- рополленинового) слоя. Пектиновый слой и сеть местами (обычно на концах клеток и вдоль гребней) прерываются округлыми отверстиями, окаймленными подпорками, которые сливаются боками. Со дна отвер- Лий отходят короткие, тоже трубчатые подпорки, от которых, возможно, берут начало длинные щетинки, пучком выходящие из отверстий. Щетинки имеют белковую природу, обнаруживают сложную микроструктуру и способствуют лучшей плавучести ценобиев: если их удалить центрифугированием, ценобии быстро оседают на дно сосуда. Видимые в оптический микроскоп гребни и шипы также образованы подпорками Рис. 92. Схема строения стенки в ценобий 5сепейевти^.
цм — цитоплазматическая мембрана, ц — целлюлозный внутренний слон стенки, сп — средник епоро- поллениновый слой, пел — пектиновый слой, пс — пектиновая сеть, о —отверстие в пектиновом слое, п — подпорки, на которых покоится сеть, ер — гребень, состоящий из более длинных подпорок, ш - шип, образованный самыми длинными подпорками, кп — короткие подпорки, поднимающиеся со диа отверстий, щ — щетинки, выходящие из отверстий разной длины. Самые длинные подпорки составляют шипы (рис. 92), При формировании клеточной стенки вокруг развивающихся автоспор первым, как и у хлореллы, откладывается- средний слой в виде трехслойных дисков, которые растут и, смыкаясь, полностью окружают клетку. Затем начинается образование внутреннего целлюлозного и наружного пектинового слоев. В последние годы появились сообщения о существовании у Зсепейезтиз оЬИцииз изогамного полового процесса: при снижении в среде содержания азота вегетативные клетки превращаются в гаметангии, в которых развиваются по восемь гамет. На гаметогенез влияют также температура и освещение. Интерес представляет род протосифон (Рго(оз1- рНоп, рис. 93) — связующее звено между хлорококко- выми и бриопсидовыми (сифоновыми) водорослями. Протосифон встречается на сырой земле и распрост- Рис. 93. Рго~ ранен повсеместно, особенно в тропиках. Многоядер- (оырНоп ная клетка, более или менее шаровидная, несущая в постенной цитоплазме сетчато-продырявленный хроматофор со многими пиреноидами, переходит в трубковидный бесцветный неразветвлен- ный ризоид. В неблагоприятных условиях наблюдается половой процесс: гаметы развиваются из вегетативных клеток, а также и из толстостенных апланоспор. * * * Хлорококковые — почти исключительно пресноводные организмы, особенно богато представленные в планктоне. Отростки клеточных стенок у многих видов педиаструма, сценедесмуса, шипы, щетинки, характерные для голенкинии, микрактиниума и др., увеличивая трение о воду, поддёрживают водоросль во взвешенном состоянии, служат приспособлениями к планктонному образу жизни. Подобно многим вольвоксов'ым, хлорококковые обнаруживают ясно выраженную тенденцию к сапротрофности. Было показано, что многие виды, хотя и могут расти в чисто минеральных средах, гораздо лучше развиваются при добавлении в среду органических веществ, особенно глюкозы. В последнем случае клетки, во всех остальных отношениях совершенно здоровые, нередко теряют окраску, которая может вновь появиться при исчерпании в среде глюкозы или при перенесении клеток в чисто минеральную среду. Миксотрофность многих хлорококковых и потеря зеленой окраски в присутствии органических веществ в среде, несомненно, благоприятствуют возникновению постоянно бесцветных сапротрофных форм. Среди хлорококковых, так же как среди вольвоксовых, можно наблюдать параллельные формы, различающиеся только по присутствию или отсутствию хлорофилла. Так, род прото- тека (РгоШНеса), встречающийся в соке, вытекающем из поврежденных стволов деревьев, отличается от хлореллы только отсутствием хлорофилла. Род гиалорафидиум (Нуа1огарНШит) — бесцветная форма, параллельная анкистродесмусу. Описаны виды харациума, лишенные окраски. Способность многих хлорококковых к питанию органическими веществами послужила, ' по-видимому, основой для сожительства с другими организмами. Ряд форм приспособился к эндофитизму. Примером их может служить упомянутый хлорохитриум. Некоторые хлорококковые вступают в сожительство с грибами, входя в состав лишайников в качестве гонидий. Другие (зоохлореллы) встречаются внутри организма животных: в инфузориях, гидре и других кишечнополостных, в пресноводных губках, моллюсках, червях и пр. В одной пресноводной губке была обнаружена колониальная водоросль Зсепейезтив циайпсаийа. Хлорококковые легко культивируются в искусственных условиях и Давно используются в качестве объектов физиологических и биохимических исследований, особенно при изучении фотосинтеза. Они служат и объектами массового культивирования для практического использования. В СССР работы по массовому культивированию водорослей начались в связи с потребностями рыбного хозяйства: были разработаны методы разведения живых кормов для молоди промысловых рыб. Один
В замкнутых системах хлорелла может применяться не только для питания человека, но и для регенерации воздуха, используя в процессе- фотосинтеза углекислоту, выдыхаемую человеком, и выделяя необходимый для его дыхания кислород. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 722; Нарушение авторского права страницы





 Рис. 85. НуЛгоНЫуоп геИсиШит. А — гамета с полярной шапочкой (/); Б — гамета без полярной шапочки; В—гамета с полярной шапочкой дает оплодотворяющую трубку (2); Г — соединение гамет посредством оплодотворяющей трубки
Рис. 85. НуЛгоНЫуоп геИсиШит. А — гамета с полярной шапочкой (/); Б — гамета без полярной шапочки; В—гамета с полярной шапочкой дает оплодотворяющую трубку (2); Г — соединение гамет посредством оплодотворяющей трубки
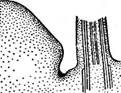

 Рис. 86. Ценобий Ре(Иа$1гит:
1 — вегетативные клетки. 2 — клетки с зооспорами. 3 — пустые клетки после выхода зооспор
Рис. 86. Ценобий Ре(Иа$1гит:
1 — вегетативные клетки. 2 — клетки с зооспорами. 3 — пустые клетки после выхода зооспор

 ■ Рис. 87. СШогеНа. А — вегетативная клетка; Б, В — образование и выход автоспор
■ Рис. 87. СШогеНа. А — вегетативная клетка; Б, В — образование и выход автоспор

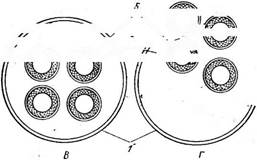
 Рис.90. Оогамия у хлорококковых. А— В— Оокпкша зоШагьа: А — вегетативная клетка; Б — сперматозоид, прикрепившийся к яйцеклетке; В — молодая, еще голая ©оспора после выхода из шиповатой оболочки оогония; Г — Ж — М1сгас1Шит ривШит". / —внешний вид колонии; Д — образование сперматозоидов; Е — слияние сперматозоида
Рис.90. Оогамия у хлорококковых. А— В— Оокпкша зоШагьа: А — вегетативная клетка; Б — сперматозоид, прикрепившийся к яйцеклетке; В — молодая, еще голая ©оспора после выхода из шиповатой оболочки оогония; Г — Ж — М1сгас1Шит ривШит". / —внешний вид колонии; Д — образование сперматозоидов; Е — слияние сперматозоида



