
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Основные физиологические понятияСтр 1 из 15Следующая ⇒
Лекция 1 Введение в курс Предмет и задачи физиологии Физиология (от греческих слов: физис - природа, логос - учение, наука) наука о функциях как организма в целом, так и отдельных составляющих его систем: органов, тканей, клеток; процессах и механизмах, обуславливающих возможность реализации и регуляции этих функций и обеспечивающих жизнедеятельность человека и животных в их взаимодействии с окружающей средой. Задача физиологии заключается в установлении физических и химических основ процессов жизнедеятельности на всех уровнях организации живых систем от субклеточного до уровня целого организма. О связи со сложностью предмета изучения физиология подразделяется на несколько разделов. К ним относятся: общая и клеточная физиология, задачей которой является изучение закономерностей основных жизненных процессов и механизмов характерных для всех функциональных и структурных систем организма и частная физиология, в свою очередь подразделяющаяся на физиологию отдельных органов: сердца, почек, печени и т.д.; физиологию функциональных систем: дыхания, кровообращения, пищеварения, эндокринной системы, физиологию развития, нейрофизиологию и физиологию поведения, сенсорную физиологию. Существует также прикладная физиология: спорта, труда, питания, космическая и подводная физиология. Каково же практическое значение физиологии? Почему так важно знать, как работает организм? Наиболее очевидная причина заключается в том, что физиология, и особенно учение о функционировании организма человека, служит научной основой медицины и фармацевтической химии. На всех прежних этапах развития цивилизации подход к лечению болезней был почти целиком эмпирический, т.е. сводился к методу проб и ошибок. В результате, постепенно было подмечено, что при тех или иных болезнях помогают определенные средства - будь то химические или растительные препараты, минеральная вода и горячие ванны, акупунктура или даже методы психологического воздействия, которые широко использовались в народной медицине. Лечебный эффект многих современных препаратов был сначала обнаружен чисто эмпирически (например, нитроглицерин, использовался Нобелем для производства динамита, при этом вызывал у рабочих сильное падение артериального давления, часто приводившее к летальному исходу), а главные механизмы, лежащие в основе целебного действия, целого ряда из них пока так и остаются неизвестными. Однако по мере развития и углубления нашего понимания механизмов функционирования и патологии органов и тканей, снижается роль чисто эмпирических находок и появляется возможность разрабатывать эффективные, научно обоснованные способы лечения заболеваний человека, целенаправленно создавать новые лекарственные препараты. Поэтому студентам, изучающим фармацевтическую химию, необходимо знать современные представления о механизмах фармакологического и токсического воздействия на организм и уметь применять полученные знания в будущей производственной деятельности. Основные физиологические понятия Каждая научная дисциплина имеет собственный понятийный аппарат. Естественно, что и в физиологии, науке сформировавшейся еще в XIX веке, существуют собственные понятия и термины. Ниже приведены наиболее общие из них. Другие, термины и понятия, имеющие более узкое значение, будут рассмотрены в ходе изложения соответствующих разделов физиологии на последующих лекциях. Основные физиологические понятия. Функция - специфическая деятельность системы или органа. Например, функциями желудочно-кишечного тракта являются моторная, секреторная, всасывательная; функцией системы дыхания - обмен СО2 и кислорода; функцией системы кровообращения - движение крови по сосудам; функцией миокарда - сокращение и расслабление, и т.д. Процесс - совокупность последовательных действий или состояний, направленных на достижение определенного результата. Механизм - способ регулирования процесса или функции. ‚ физиологии принято выделять нервный и гуморальный (т.е. посредством выделения гормонов или других химических биологически активных веществ) механизмы регуляции. Регуляция - изменение интенсивности и направленности функции (процесса) с целью обеспечения оптимальной деятельности органов и систем. Система в физиологии это совокупность органов, тканей или других структурных образований, связанных общей функцией. Например, сердечно-сосудистая система обеспечивает с помощью сердца и сосудов доставку тканям питательных, регуляторных, защитных веществ и кислорода, а также отвод продуктов обмена; экспортная (секреторная) система клетки обеспечивает с помощью эндоплазматического ретикулума и аппарата Гольджи синтез и транспортировку из клеток гормонов, липопротеидов и других секретов. Секрет - специфический продукт жизнедеятельности клетки, выполняющий определенную функцию и выделяющийся во внутреннюю среду организма. Процесс выработки и выделения секрета называется секрецией. По составу секрет делят на белковый (серозный), слизистый (мукоидный), смешанный и липидный. Раздражитель - факторы внешней и внутренней среды или их изменение, которые оказывают на органы и ткани влияния, приводящие к изменению активности последних. Различают раздражители механические, электрические, химические, температурные, звуковые и т.д. Раздражитель может быть по величине пороговым, т.е. оказывать минимальное эффективное воздействие; максимальным, дальнейшее увеличение силы которого не приводит к увеличению соответствующей реакции; сверхсильным, действие которого может оказывать повреждающий или болевой эффект или приводить к неадекватным ощущениям. Раздражение - воздействие на живую ткань внешних и внутренних раздражителей. Реакция - изменение (усиление или ослабление) деятельности организма или его составляющих в ответ на раздражение. Рефлекторная реакция - ответное действие (или процесс) в организме (системе, органе, ткани, клетке), вызванное рефлексом. Рефлекс - возникновение или изменение функциональной активности органов, тканей или целостного организма, осуществляемое при участии центральной нервной системы в ответ на раздражение нервных окончаний (рецепторов). Возбудимость - способность живых клеток воспринимать изменения внешней среды и отвечать на эти изменения реакцией возбуждения. Чем ниже пороговая сила раздражителя, тем выше возбудимость, и наоборот. Возбуждение - активный физиологический процесс, которым некоторые живые клетки (нервные, мышечные, железистые) отвечают на внешнее воздействие. Возбудимые ткани - ткани, способные в ответ на внешнее воздействие, превышающее порог возбуждения, переходить из состояния физиологического покоя в состояние возбуждения. ‚ принципе, все живые клетки обладают той или иной степенью возбудимости, но в физиологии к этим тканям принято относить нервную, мышечную, железистую. Физиология клетки Клеточные мембраны. Лекция 2 Трансмембранный транспорт. Мембранный потенциал. Разность электрических потенциалов (в вольтах или мв) между жидкостью, находящейся по одну сторону мембраны и жидкостью по другую ее сторону называется мембранным потенциалом (МП) и обозначается Vм. Величина МП живых клеток составляет обычно от -30 до -100 мв и вся эта разность потенциалов создается в областях непосредственно прилегающих с обоих сторон к клеточной мембране. Уменьшение величины МП называют деполяризацией, увеличение - гиперполяризацией, восстановление исходного значения после деполяризации - реполяризация. Мембранный потенциал существует во всех клетках, но в возбудимых тканях (нервных, мышечных, железистых), мембранный потенциал или как его еще называют в этих тканях, мембранный потенциал покоя, играет ключевую роль в реализации их физиологических функций. Мембранный потенциал обусловлен двумя основными свойствами всех эукариотических клеток: 1) асимметричным распределением ионов между вне- и внутриклеточной жидкостью, поддерживаемым метаболическими процессами; 2) Избирательной проницаемостью ионных каналов клеточных мембран. Чтобы уяснить себе как возникает МП представим себе, что некий сосуд разделен на два отсека мембраной, проницаемой только для ионов калия. Пусть в первом отсеке содержится 0, 1 М, а во втором 0, 01 М раствор КСl. Поскольку концентрация ионов калия (К+) в первом отсеке в 10 раз выше, чем во втором, то в начальный момент на каждые 10 ионов К+ диффундирующих из отсека 1 во второй будет приходится один ион диффундирующий в обратном направлении. Так как анионы хлора (Сl-) не могут переходить через мембрану вместе с катионами калия, то во втором отсеке будет образовываться избыток положительно заряженных ионов и, напротив, в отсеке 1 окажется избыток ионов Сl-. В результате возникает трансмембранная разность потенциалов, препятствующая дальнейшей диффузии К+ во второй отсек, поскольку для этого им нужно преодолеть притяжение отрицательных ионов Сl-, в момент вхождения в мембрану со стороны отсека 1 и отталкивание одноименных ионов на выходе из мембраны в отсек 2. Таким образом, на каждый ион К+, проходящий через мембрану в этот момент действуют две силы - химический градиент концентраций (или химическая разность потенциалов), способствующая переходу ионов калия из первого отсека во второй, и электрическая разность потенциалов, заставляющая ионы К+ двигаться в обратном направлении. После того как эти две силы уравновесятся, количество ионов К+ перемещающееся из отсека 1 в отсек 2 и обратно сравняется, установится электрохимическое равновесие. Соответствующая такому состоянию трансмембранная разность потенциалов называется равновесным потенциалом, в данном конкретном случае равновесным потенциалом для ионов калия (Ек). В конце 19 века Вальтер Нернст установил, что равновесный потенциал зависит от абсолютной температуры, валентности диффундирующего иона и от отношения концентраций данного иона по разные стороны мембраны: Ex = где Ех- равновесный потенциал для иона X, R - универсальная газовая постоянная = 1, 987 кал/(моль град), T- абсолютная температура в градусах Кельвина, F - число Фарадея = 23060 кал/в, Z - заряд переносимого иона, [X]1 и [X]2 - концентрации иона в отсеках 1 и 2. Если перейти от натурального логарифма к десятичному, то для температуры 18˚ С и моновалентного иона можно записать уравнение Нернста следующим образом: Ех= 0, 058 Рассчитаем с помощью уравнения Нернста калиевый равновесный потенциал для воображаемой клетки, приняв, что внеклеточная концентрация калия [К+]н= 0, 01 М, а внутриклеточная - [К+]в = 0, 1 М: Ек= 0, 058
В данном случае, Екотрицателен, поскольку ионы калия будут выходить из гипотетичной клетки, заряжая отрицательно слой цитоплазмы, прилегающий к внутренней стороне мембраны. Поскольку в данной гипотетичной системе имеется только один диффундирующий ион, то калиевый равновесный потенциал будет равен мембранному потенциалу (Ек= Vм).
Приведенный механизм ответственен и за образование мембранного потенциала в реальных клетках, но в отличие от рассмотренной упрощенной системы, в которой через " идеальную" мембрану мог диффундировать только один ион, реальные клеточные мембраны пропускают в той или иной все неорганические ионы. Однако, чем менее мембрана проницаема для какого-либо иона, тем меньшее влияние он оказывает на МП. Учитывая это обстоятельство, Голдманом в 1943г. было предложено уравнение для расчета величины МП реальных клеток, учитывающее концентрации и относительную проницаемость через плазматическую мембрану всех диффундирующих ионов: Vм = 0, 058 Используя метод меченых изотопов, Ричард Кейнс в 1954 г. определил проницаемость клеток мышц лягушки для основных ионов. Оказалось, что проницаемость для натрия примерно в 100 раз меньше, чем для калия, а ион Сl-не вносит заметного вклада в создание МП мышечных клеток. Поэтому для мембран мышечных клеток уравнение Голдмана можно записать в следующем упрощенном виде: Vм = 0, 058 Vм = 0, 058
Исследования с применением вводимых в клетки микроэлектродов, показали, что потенциал покоя клеток скелетных мышц лягушки колеблется от -90 до -100 мв. Такое хорошее соответствие экспериментальных данных теоретическим подтверждает, что потенциал покоя определяется диффузионными потоками неорганических ионов. При этом, в реальных клетках мембранный потенциал близок к равновесному потенциалу иона, который характеризуется максимальной трансмембранной проницаемостью, а именно к равновесному потенциалу иона калия. Роль кальция Лекция 3 Основы биоэнергетики. Химическая энергия, особенности ее образования и утилизации в живых системах. Молекулярные механизмы преобразования энергии в живых клетках изучает биоэнергетика. Восстановленные органические соединения, служащие источником энергии живым организмам, окисляются до СО2 и Н2О, а высвобождающаяся энергия используется для синтеза АТФ и выполнения различных работ. Прежде чем перейти к рассмотрению конкретных биоэнергетических процессов в клетке вспомним, что в соответствии со вторым законом термодинамики энтропия любой замкнутой системы с течением времени может только возрастать. Энтропия - термодинамическая категория, характеризующая степень неупорядоченности системы. Поэтому, возрастание энтропии, по сути, есть увеличение степени неупорядоченности системы, т.е. переход от более упорядоченной структуры к менее упорядоченной и снижение доли полезной энергии (т.е. энергии, с помощью которой может быть совершена работа). Другими словами из второго закона термодинамики следует, что в любой изолированной системе степень неупорядоченности может только возрастать, а величина полезной (свободной) энергии - уменьшаться. В тоже время живые организмы, по мере того как проходят развитие от оплодотворенной яйцеклетки до состояния взрослой особи, постоянно усложняют свою структуру. Казалось бы, в живых системах второй закон термодинамики не выполняется. Однако, следует иметь ввиду, что этот закон относится к замкнутой системе, а живой организм является открытой системой, для которых характерен постоянный обмен с окружающей средой веществом и энергией и поддерживание относительно низкого уровня энтропии, за счет повышения ее в окружающей среде. Способность извлекать полезную энергию из окружающей среды является одним из тех замечательных свойств, которые отличают живые организмы от неживой природы. Потребляя растительную или животную пищу, живой организм превращает содержащиеся в ней высокоорганизованные молекулы в СО2, Н2О и низкомолекулярные азотсодержащие соединения, имеющие значительно более высокий уровень энтропии, а образующуюся при этом энергию использует, в том числе на развитие и рост собственного организма, сопровождающийся понижением энтропии. Первичным же источником энергии на земле для большинства организмов является солнце, энтропия которого постоянно повышается и только незначительная часть выделяемой при этом энергии улавливается биосферой и утилизируется растениями и некоторыми бактериями в реакциях фотосинтеза, а остальная безвозвратно теряется во Вселенной. Даже открытая недавно обильная жизнь глубоко на дне океанов в области океанических трещин на т.н. «черных курильщиках» зависит от солнца, поскольку является аэробной, а кислород берется в атмосфере и океане только благодаря процессу фотосинтеза. В процессе эволюции в организме животных и естественно человека сформировалось множество путей получения, транспорта и утилизации энергии, и в подавляющем большинстве из них для аккумуляции химической энергии используются специальные соединения – фосфагены, имеющие в своей структуре так называемую высокоэнергетическую или макроэргическую связь. Аденозин-5'-трифосфат (АТФ) представляет собой наиболее распространенный и важнейший представитель этой группы соединений. Кроме АТФ, высокоэнергетические связи имеют ряд других органических полифосфатов, например ГТФ, ЦТФ Для обозначения макроэргической связи используется символ ~. Так молекулу АТФ сокращенно обозначают А-Р-Ф~Ф~Ф (А-аденин, Р-рибоза). Некоторые бактерии даже запасают энергию в форме неорганического полиметафосфата – линейного полимера неопределенной длинны. При рН=7, 0 АТФ представляет анион с высоким зарядом. Трифосфатная группа содержит 4 ОН группы способные к ионизации. 3 из них имеют низкое значение рК и при рН=7 полностью диссоциируют, четвертая группа диссоциирует на 75%. Высокая концентрация отрицательных зарядов является важным фактором, обуславливающим высокоэнергетическую природу АТФ. Эти заряды располагаются близко друг к другу и между ними существует сильное отталкивание. При гидролитическом отщеплении концевой фосфатной группы силы отталкивания ослабевают. Образовавшиеся продукты не стремятся воссоединиться. Вторая важная особенность, обуславливающая большую отрицательную величину Δ G' гидролиза АТФ заключается в том, что оба продукта гиролиза стабилизированы за счет сопряжения. Впрочем, в клетках АТФ почти не встречается в форме свободных ионов, а чаще в виде комплекса с ионами Mg или Mn. Понятие энергии фосфатной связи, которым пользуются биохимики не следует путать с понятием энергии связи, под которой в физической химии понимают энергию необходимую для разрыва в молекуле связи между 2 атомами. Под энергией макроэргической фосфатной связи понимают разность между свободной энергией исходных веществ и свободной энергией продуктов реакции, в процессе которой фосфорилированное соединение гидролизуется с образованием неорганического фосфата. Перенос концевой фосфатной группы АТФ на воду сопровождается освобождением свободной энергии, примерно вдвое большей, чем при гидролизе обычной фосфоэфирной связи, например в аденозинмонофосфате. Свободная энергия характеризует ту долю полной энергии заключенной в веществе, с помощью которой может быть совершена работа в изотермических и изобарических условиях, т.е. при постоянной температуре и давлении, а именно такие условия характерны для биологических систем. Изменение свободной энергии измеряется в ккал/моль и обозначается Δ G' если процесс протекает в стандартных условиях (рН=7, Т=25 º С, субстраты и продукты реакции взяты в одинаковой концентрации равной 1 моль/л) и символом Δ G для реакции в конкретных условиях реальных биологических систем. В случае гидролиза АТФ, Δ G' = -7 ккал/моль, а Δ G = -11-13 ккал/моль Живые организмы используют энергию АТФ для выполнения механической работы мышц, создания концентрационных градиентов и трансмембранных электрических потенциалов. Конкретные механизмы, обеспечивающие протекание данных процессов будут изложены в последующих лекциях, а теперь рассмотрим, общие принципы, обеспечивающие возможность использования энергии АТФ для совершения химической работы. Представим себе типичный биосинтетический, т.е. анаболический процесс (анаболизм от греч. подъем те процессы синтеза, построения структурных частей клетки, противоположен катаболизму), при котором два мономера -Х и Y- должны соединиться друг с другом в реакции дегидратации (называемой также конденсацией), сопровождающейся выделением воды: Х-ОН + Y-Н Большинство таких реакций являются эндергоническими (не путать с эндотермическими) и сопровождаются возрастанием свободной энергии на величинуΔ G, а движущей силой любой химической реакции является уменьшение свободной энергии и самопроизвольно протекают только экзергонические реакции и, следовательно, почти всегда предпочтительнее обратная катаболическая реакция (называемая гидролизом), поскольку в этом случае имеет место выделение свободной энергии равное по абсолютной величине ее потреблению в реакции конденсации. Поэтому в клетке такие реакции протекают в несколько стадий, в результате которых происходит сопряжение энергетически невыгодного синтеза с сильно экзергоническиой реакцией, протекающей с участием АТФ. Механизм сопряжения реализуется следующим образом. Реакции синтеза, приведенной выше и сопровождающейся возрастанием свободной энергии (предположим Δ G' = 3 ккал/моль), предшествует фосфорилирование одного из субстратов молекулой АТФ. А-Р-Ф~Ф~Ф + ХОН → А-Р-Ф~Ф + Х-О-Ф - Δ G = 4 ккал/моль Δ G' этой реакции значительно меньше (4 ккал/моль), чем реакции гидролиза АТФ и переноса фосфатной группы на воду (примерно 11 ккал/моль). Остальная энергия (7 ккал/моль) аккумулируется в молекуле активированного субстрата, который вовлекается в последующую синтетическую реакцию. Однако, теперь свободная энергия исходных веществ выше, чем свободная энергия продуктов реакции (7 - 3 = 4 ккал/моль) и, реакция протекает слева направо, т.е. в направлении синтеза, сопровождаясь уменьшением свободной энергии: Х-О-Ф + Y-Н → X-Y + Фн - Δ G = ( 4 ккал/моль) Таким образом, за счет накопленной в молекуле АТФ энергии эндергоническая реакция замещается двумя сопряженными экзергоническими реакциями. Другой путь использования АТФ для совершения химической работы реализуется через последовательно сопряженные или как их называют тандемные реакции. В этом случае эндергоническая реакция протекает благодаря следующей за ней экзергонической реакции. Возможность протекания тандемных реакций обусловлена тем обстоятельством, что увеличение свободной энергии не исключает протекание химической реакции, а лишь указывает на то, что величина константы равновесия такой реакции меньше единицы, т.е. если имеет место реакция:
А + B
то Кeq = и, следовательно, равновесие реакции смещено влево и это смещение тем более выражено, чем больше по абсолютной величине Δ G'. Однако если за реакцией 1 следует реакция 2 С +X и для этой реакции величина Δ G' сильно отрицательна, то промежуточный продукт реакции С будет потребляться, смещая при этом равновесие реакции 1 вправо. В том случае если суммарная свободная энергия двух реакций будет величиной отрицательной они будут протекать в направлении потребления субстратов А и В и образования продукта СХ. Теперь рассмотрим где в клетке локализованы механизмы получения энергии и перевода фосфагенов из " незаряженной" формы (например АДФ) в " заряженную" (АТФ) и как они функционируют. Цикл трикарбоновых кислот В большинстве клеток и тканей аэробных организмов, образующаяся в результате гликолиза пировиноградная кислота поступает в матрикс митохондрий, где она декарбоксилируется, т.е. от нее отщепляется молекула углекислого газа, а оставшаяся двухуглеродная ацетильная группа соединяется с коферментом А (СоА), образуя ацетил-СоА. Энергия, выделяющаяся в результате этой реакции, расходуется на перевод специальной молекулы - переносчика водорода из окисленной в восстановленную форму. Переносчик водорода называется никотинамидадениндинуклеотид и обозначается сокращенно NAD (НАД) или NAD+(окисленная форма) и NADH (НАДН) или NADH + H+(восстановленная форма). Таким образом, молекулы NAD+ и сходного с ним по структуре и функции флавинадениндинуклеотида, сокращенно FAD, подобно фосфагенам способны запасать и переносить энергию, но не в виде макроэргической фосфатной связи, а в виде активированных атомов водорода. Образовавшийся из пировиноградной кислоты ацетил-СоА подвергается дальнейшему окислению в цикле трикарбоновых кислот. В результате первой реакции цикла (Рис. ) из ацетил-С и щавелевоуксусной кислоты (в водной среде существует в виде иона - оксалоацетата) содержащей 4 атома углерода при участии фермента цитратсинтазы образуется трикарбоновая, шестиуглеродная лимонная кислота (цитрат). Поэтому цикл трикарбоновых кислот называют иногда цикл лимонной кислоты. Затем лимонная кислота превращается в изолимонную (изоцитрат). В результате следующей, реакции декарбоксилирования шестиуглеродная молекула изолимонной кислоты превращается в пятиуглеродную α -оксоглутаровую кислоту (α -оксоглутарат) и происходит восстановление NAD+ в NADH. Далее α -оксоглутаровая кислота декарбоксилируется до янтарной (сукцинат). При этом выделяющаяся энергия не рассеивается в виде тепла, а затрачивается на восстановление еще одной молекулы NAD+ в NADH и образование промежуточного продукта с высокоэнергетической фосфатной связью, который обеспечивает фосфорилирование гуанозиндифосфата (GDP)в гуанозинтрифосфат (GTP).
В результате трех следующих реакций янтарная кислота последовательно превращается в фумаровую кислоту (фумарат), яблочную кислоту (малат) и щавелевоуксусную и восстановливается по молекуле FAD и NAD. С точки зрения биоэнергетики при каждом обороте цикла удаляются 2 атома углерода вошедшего в цикл как ацетил-СоА в виде СО2 и генерируется 8[H] + 2[H] на входе – это активированный водород -источник энергии. Таким образом, ферментативные реакции цикла трикарбоновых кислот протекают так, что субстрат первой реакции - щавелевоуксусная кислота является и продуктом последней восьмой реакции, т.е. регенерируется после каждого оборота цикла и процесс запускается вновь. В результате элементарного цикла два углеродных атома, имевшихся в ацетил-СоА, превращаются в СО2. Циклический характер последовательности реакции окисления ацетил-СоА и ее основные этапы установил в 1937 году английский биохимик немецкого происхождения, лауреат Нобелевский премии (1953) Ханс Кребс. Поэтому процесс окисления ацетил-СоА имеет еще одно название - цикл Кребса. Благодаря тому, что окисление совершается постепенно, через серию относительно небольших изменений свободной энергии, выделяющаяся энергия не рассеивается в виде бесполезного тепла, а затрачивается на восстановление трех молекул NAD в NADH и одной молекулы FAD в FADH и образование высокоэнергетической фосфатной связи. Эта связь образуется в результате фосфорилирования гуанозиндифосфата в гуанозинтрифосфат. Последний легко обменивается фосфатной группой с АДФ с образованием АТФ. Следует иметь в виду, что источником ацетил-CoA для окисления в ЦТК, наряду с гликолизом являются и другие биохимические процессы протекающие в клетках, в частности окисление аминокислот и жирных кислот (а также этанол). Четыре молекулы NADH, включая и образовавшуюся при окислении пирувата в ацетил-СоА, и молекула FADH, поступают в дыхательную цепь, где переносимые ими атомы водорода окисляются молекулярным кислородом до воды. Выделяемая при этом энергия затрачивается на синтез АТФ из АДФ и неорганического фосфата в результате сложнейшего, во многом неясного процесса, получившего название окислительного фосфорилирования. Лекция 4 Лекция 5. Лекция 6. Лизосомы: структура и пути образования в клетке, классификация. Лизосомы представляют собой гетерогенную (разнородную) группу цитоплазматических вакуолеподобных структур размером 1-3 мкм, отличительной особенностью которых является наличие в них кислой среды и большого количества различных гидролаз - ферментов способных расщеплять (гидролизовать) основные типы макромолекул. Присутствие в лизосомах гидролаз определяет их основную функцию в клетке - расщепление макромолекул и более крупных образований как поступающих в клетку из внеклеточного пространства так и имеющих внутриклеточное происхождение. Интересно, что у растений нет лизосом. Известны три возможных пути образования лизосом в клетке. В каждом случае образуются морфологически различные образования, расщепляющие материал из различных источников.
В первом случае расщепляемый материал - белки, полинуклеотиды или полисахариды - попадает в клетку путем эндоцитоза. В ходе этого процесса молекулы, имеющие достаточно крупные размеры и неспособные проникать через мембраны, постепенно окружаются небольшим участком плазмалеммы, который сначала впячивается (инвагинируется), а затем отщепляется вовнутрь клетки, образуя пузырек, содержащий захваченный клеткой материал. Пузырьки образующиеся в результате эндоцитоза получили название эндосомы. По мере движения эндосомы от клеточной мембраны во внутрь клетки она многократно взаимодействует с транспортными пузырьками доставляющими от транс-поверхности аппарата Гольджи гидролитические ферменты и мембранные белки, превращаясь в эндолизосому. Процесс образования и трансформации эндосомы длится около 15 мин и сопровождается закислением внутренней среды, благодаря закачиванию ионов Н+из цитозоля во внутрь эндосомы АТФ-зависимым протонным насосом, функционирующим подобно АТФ -азе внутренней мембраны митохондрий. Второй путь формирования лизосом называется аутофагией. В процессе аутофагии происходит разрушение отработанных частей самой клетки. Известно, например, что в клетках печени среднее время жизни одной митохондрии составляет около 10 дней, после чего она должна быть утилизирована в лизосомах. На электронных микрофотографиях нормальных клеток можно увидеть лизосомы содержащие митохондрии на разных стадиях деградации. Выше уже отмечалось, что путем аутофагии из клеток печени удаляется избыток гладкого ЭР, после прекращения поступления и выведения из организма ксенобиотиков - индукторов. Процесс аутофагии, по-видимому, начинается с окружения органеллы мембранами, поставляемыми из ЭР, в результате чего образуется аутофагосома. Затем, полагают, что аутофагосома сливается с эндолизосомой, образуя аутофаголизосому, в которой и происходит процесс деградации фрагмента ЭР или другой органеллы. Третий путь формирования лизосом имеется только у клеток, специализированных для фагоцитоза больших частиц и микроорганизмов. Такие клетки-фагоциты, а к ним относятся клетки крови - нейтрофилы и моноциты, могут поглощать из внеклеточного пространства крупные объекты, образуя фагосомы. Далее фагосома превращается в фаголизосому тем же путем, что и аутофагосома, т.е. сливаясь с эндолизосомой. Эндосомы, аутофагосомы и фагосомы часто называют общим термином - прелизосомы, а эндолизосомы, аутофаголизосомы и фаголизосомы термином лизосомы. В зрелых лизосомах происходит деградация поглощенного материала до отдельных молекул, например аминокислот, которые поступают в цитозоль и вовлекаются в последующие биохимические превращения. Фрагменты собственной плазматической мембраны не подвергаются воздействию гидролаз и возвращается обратно в плазмалемму с помощью транспортных пузырьков, еще до окончательного формирования лизосомы. Неперевариваемые продукты остаются и накапливаются в лизосомах, которые теряют гидролитические ферменты и превращаются в постлизосомы или остаточные тельца. С возрастом, в клетках человека и животных увеличивается количество остаточных телец, содержащих большое количество липофусцина или пигмента старения. Липофусцинпредставляет собой биополимеры различной природы, неподдающиеся дальнейшему расщеплению поскольку химические связи между отдельными мономерами образовались не в нормальных биохимических реакциях, а в результате спонтанных окислительных процессов, главным образом свободнорадикальных. Различные заболевания, воздействие радиации и других негативных факторов внешней среды ускоряют процесс накопления пигмента старения. Клеточное ядро и геном Лекция 7 Лекция 1 Введение в курс Предмет и задачи физиологии Физиология (от греческих слов: физис - природа, логос - учение, наука) наука о функциях как организма в целом, так и отдельных составляющих его систем: органов, тканей, клеток; процессах и механизмах, обуславливающих возможность реализации и регуляции этих функций и обеспечивающих жизнедеятельность человека и животных в их взаимодействии с окружающей средой. Задача физиологии заключается в установлении физических и химических основ процессов жизнедеятельности на всех уровнях организации живых систем от субклеточного до уровня целого организма. О связи со сложностью предмета изучения физиология подразделяется на несколько разделов. К ним относятся: общая и клеточная физиология, задачей которой является изучение закономерностей основных жизненных процессов и механизмов характерных для всех функциональных и структурных систем организма и частная физиология, в свою очередь подразделяющаяся на физиологию отдельных органов: сердца, почек, печени и т.д.; физиологию функциональных систем: дыхания, кровообращения, пищеварения, эндокринной системы, физиологию развития, нейрофизиологию и физиологию поведения, сенсорную физиологию. Существует также прикладная физиология: спорта, труда, питания, космическая и подводная физиология. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 3624; Нарушение авторского права страницы

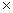 lg
lg 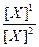
 = 0, 058
= 0, 058  =0, 058
=0, 058 


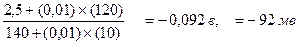

 X-Y + Н2О + Δ G (3 ккал/моль)
X-Y + Н2О + Δ G (3 ккал/моль)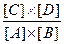 < 1
< 1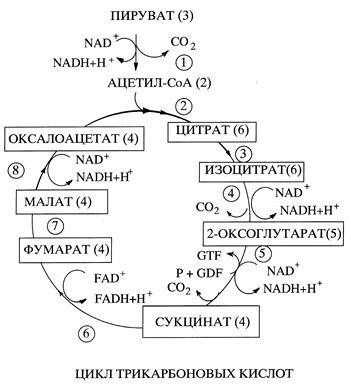
 Рис. 6 Пути образования лизосом в клетке
Рис. 6 Пути образования лизосом в клетке