Номенклатура и характеристика рестриктаз
В 1973 году Смит и Натанс предложили номенклатуру рестриктаз, включающую следующие пункты:
1. Аббревиатура названия каждого фермента является производной от бинарного названия микроорганизма, содержащего данную метилазно-рестриктазную систему. Составляют по правилу: к первой прописной букве названия рода добавляют две первые строчные буквы вида.
Streptomycesalbus - Sal, Escherichiacoli-Eco
2. В случае необходимости добавляют обозначение серотипа или штамма, например, Есо B.
3. Различные системы рестрикции - модификации, кодируемые одной бактериальной клеткой, обозначают римскими цифрами: Hind II, Hind I, Hind III (Haemophilusinfluenzae).
4. Рестриктазы обозначают буквой R (R Hind III), метилазы - М (М Hind III).
Открытие новых рестриктаз заставило Робертса в 1978 году внести дополнения в систему рациональных обозначений ферментов: если сокращенное название совпадает для нескольких ферментов, то 2 первые буквы аббревиатуры остаются неизменными, а третья берется из последующих букв видового названия:
Haemophilusparainfluenzae - Hpa I
Haemophilusparahaemolyticus - Hph I.
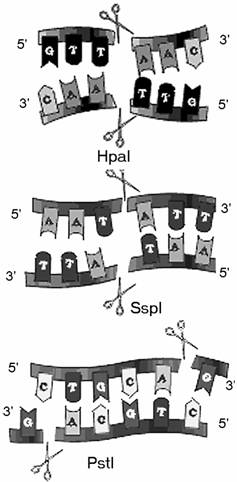 Рестриктазы по-разному расщепляют ДНК (рис. 36). Рестриктазы по-разному расщепляют ДНК (рис. 36).
Одни вносят разрывы по оси симметрии узнаваемой последовательности (Hpa I, Ssp I).
Другие - со сдвигом, со " ступенькой" (Pst I).
В первом случае образуются так называемые " тупые" концы, а во втором - " липкие", то есть фрагменты имеют на своих концах однонитевые взаимно комплементарные участки длиной в четыре нуклеотида. Такие фрагменты особенно удобны для создания рекомбинантных ДНК.
Читать дальше ► механизм действия рестриктаз
|
| | Генная инженерия, методы молекулярной биологии и генетики, связанные с целенаправленным конструированием новых, не существующих в природе сочетаний генов. Возникла в начале 70-х гг. 20 в. Основана на извлечении из клеток какой-либо организма гена (кодирующего нужный продукт) или группы генов, на соединении их со специальными молекулами ДНК (так называемыми векторами), способными проникать в клетки другого организма (главным образом микроорганизмов) и размножаться в них. Наряду с клеточной инженерией лежит в основе современной биотехнологии. Открывает новые пути решения некоторых проблем генетики, медицины, сельского хозяйства. С помощью генной инженерии был получен ряд биологически активных соединений - инсулин, интерферон и др.
| |
| | В основе Г. и. лежат достижения молекулярной биологии и прежде всего установление универсальности генетического кода (у всех организмов включение одних и тех же аминокислот в строящуюся полипептидную цепь белка кодируется одними и теми же последовательностями трех нуклеотидов в цепи ДНК). Возможность получения и направленного использования фрагментов нуклеиновых кислот, выделения отдельных участков полинуклеотидной цепи с точностью до одного нуклеотида и осуществления invitro синтеза нуклеиновых кислот с новыми сочетаниями нуклеотидных звеньев, появилась у генетической инженерии благодаря успехам энзимологии. Изменение наследственных свойств организма с помощью Г. и. состоит в конструировании из различных фрагментов ДНК нового генетического материала, введении этого материала в организм реципиента, создании условий для функционирования введенного генетического материала и его стабильного наследования. Гены или фрагменты ДНК могут быть получены путем химического синтеза. Однако этот процесс трудоемок и требует знания последовательности нуклеотидов, составляющих < < Ген> >. Более эффективен метод синтеза структурного гена на информационной РНК (иРНК) как на матрице с помощью фермента РНК-зависимой ДНК-полимеразы (обратной транскриптазы). Однако структурные гены в организме составляют функциональный комплекс с регуляторными элементами, нуклеотидные последовательности которых не воспроизводятся в молекуле иРНК. Поэтому указанный способ не позволяет осуществить синтез структурной и регуляторной области гена в совокупности, т.е. функционирующего гена в целом. Методы получения целого функционирующего гена были впервые разработаны на бактериальных клетках. Их основой является способность некоторых плазмид — небольших молекул ДНК, способных реплицироваться независимо от бактериальной хромосомы, — внедряться в хромосому бактериальной клетки, а затем спонтанно или под воздействием индуцирующих агентов, например УФ-излучения, снова переходить в цитоплазму, захватывая при этом прилегающие гены хромосомы клетки-хозяина. В цитоплазме эти гены реплицируются (размножаются) в составе захвативших их плазмид. Фрагмент генетического материала хромосомы бактериальной клетки может быть отделен от плазмиды. Использование плазмид позволяет получать в изолированном виде практически любые бактериальные гены (см. < < Плазмиды> > ). Успеху Г. и. способствовала разработка техники объединения генов, выделенных из различных источников, в одну молекулу ДНК. Решающим в конструировании таких гибридных, или рекомбинантных, молекул invitro явилось обнаружение и получение особых ферментов — рестриктаз, которые сейчас служат основным инструментом Г. и. Рестриктаза «разрезает» молекулу ДНК в участке (сайте), строго определенном для каждой конкретной рестриктазы. К середине 80-х гг. 20 в. в мировой коллекции рестриктаз насчитывалось более 400 ферментов, «узнающих» около 100 различных по структуре участков в молекулах ДНК. С помощью рестриктаз стало возможным выделение практически любого гена в виде одного или нескольких фрагментов ДНК. Появилась возможность снабжать синтезированные, «сконструированные» и природные гены различными регуляторными нуклеотидными последовательностями, заменять, вставлять, удалять нуклеотиды в строго заданных участках гена, укорачивать или достраивать его. Для создания гибридных (рекомбинантных) молекул ДНК полученные фрагменты соединяют с ДНК вектора. С этой целью используют фермент, соединяющий фрагменты ДНК. Функциональную полноценность гибридной молекулы ДНК определяют с помощью ее переноса в клетку-реципиент по последующему размножению в ней (амплификации) и функционированию. Перенос чужеродного генетического материала в реципиентные клетки и его размножение в них обеспечивает векторная часть гибридной молекулы, а синтез специфического белка — встроенный фрагмент. Прогресс Г. и. во многом обусловлен получением новых специализированных векторов. Для клонирования (размножения) сравнительно небольших фрагментов ДНК длиной до 10 тыс. пар нуклеотидов используют плазмидные векторы. Фрагменты ДНК длиной до 10—25 тыс. пар нуклеотидов клонируют с помощью векторов, полученных на основе фага лямбда. Гибридный геном такого фага, содержащий фрагмент чужой ДНК, искусственным путем «упаковывают» в белковую оболочку и этим реконструированным фагом заражают бактерии. Гибридный фаг приразмножении лизирует клетку, образуя несколько тысяч копий, которые выделяются в культуральную среду. Для клонирования фрагментов ДНК длиной до 35—45 тыс. пар нуклеотидов используют космидные векторы, представляющие собой гибрид фага лямбда и плазмиды. Космиды содержат так называемые COS-последовательности ДНК фага, необходимые для упаковки геномов фага в белковую оболочку, и участок ДНК плазмиды, позволяющий космидным векторам размножаться в бактериях, не лизируя их.Для клонирования небольших фрагментов ДНК длиной до 300—400 пар нуклеотидов в качестве векторов используют производные геномов фагов с однонитевой молекулой ДНК. Вектор, несущий чужеродную ДНК, вводят в бактериальную клетку, где эти гибридные фаги размножаются, не лизируя клетку-хозяина, и «отпочковываются» в культуральную среду как вирусные частицы с однонитевой молекулой ДНК. Сравнивая структуру фрагментов геномной ДНК и соответствующей клонированной ДНК-копии (кДНК), получают важную информацию об организации генетического материала, а в случае наследственных болезней — о характере аномалий в клонированном генетическом материале, следствием которых и является это заболевание.Созданы полные библиотеки геновмногих микроорганизмов, растений и животных (вплоть до млекопитающих и человека). Клонировано и в той или иной мере изучено около 1000 генов и других последовательностей нуклеотидов ДНК человека. Возможности Г. и. не ограничиваются клонированием гена и получением большого числа его копий. Часто необходимо обеспечить экспрессию гена в клетке, т.е. реализовать заключенную в нем генетическую информацию. Если вводимый в бактериальную клетку ген получен из бактерий той же (или близкой) видовой принадлежности, то достаточно выделить ген с его собственными регуляторными элементами, контролирующими экспрессию. Однако, если не считать нескольких исключений, регуляторные нуклеотидные последовательности эволюционно далеких друг от друга организмов не являются взаимозаменяемыми. Поэтому, чтобы добиться, например, экспрессии гена высших организмов в клетках Е, coli, у него удаляют регуляторную область, а структурную часть такого гена присоединяют (на определенном расстоянии) к регуляторной области бактериального гена. Существенный прогресс в разработке этой методики был достигнут после открытия фермента нуклеазы BAL31, которая обладает уникальным свойством удалять с конца фрагмента ДНК «лишние» последовательности нуклеотидов любой протяженности. В настоящее время структурную и регуляторную области выделяют порознь с помощью тех рестриктаз, участки «узнавания» которых расположены наиболее удачно на полинуклеотидной цепи. Затем убирают «лишние» нуклеотидные последовательности и соединяют структурную область гена высшего организма (эукариотического гена) с регуляторной областью бактериального гена. Таким путем удается добиться не только экспрессии генов эукариотов в бактериальных клетках, но и, наоборот, бактериальных генов в клетках высших и низших эукариотов. В качестве реципиентов эффективно используют не только бактериальные клетки, но и клетки высших организмов. С помощью методов Г. и. были обнаружены отклонения в строении определенных участков генов человека, которые являются причиной наследственных болезней. Чаще всего таким методом служит так называемыйблот-анализ. Выделенную клеточную ДНК подвергают расщеплению рестриктазой, полученные фрагменты разделяют по величине с помощью электрофореза. Однотипные фрагменты ДНК инкубируют с ранее клонированным геном (или его частью) либо с полученной путем химического синтеза последовательностью нуклеотидов, содержащих радиоактивную метку. Меченая ДНК связывается только с теми фрагментами анализируемой клеточной ДНК, которые имеют комплементарные ей последовательности нуклеотидов (см. < < Нуклеиновые кислоты> > ). Изменение распределения и количества фиксированной метки по сравнению с нормой позволяет судить о перестройках в анализируемом гене или в близлежащих к нему последовательностях нуклеотидов. Участки «узнавания» определенных рестриктаз в молекуле ДНК располагаются неравномерно, поэтому при расщеплении этими ферментами молекула ДНК распадается на ряд фрагментов различной длины (так называемые рестрикционные фрагменты). Перестройка структуры ДНК, в результате которой исчезают имевшиеся или появляются новые участки «узнавания».приводит к изменению набора этих фрагментов, т.е. к появлению полиморфизма длин рестрикционных фрагментов (ПДРФ). Перестройки в молекуле ДНК могут вызывать или не вызывать изменения в процессе синтеза или в структуре кодируемого белка; перестроек, не вызывающих изменений, большинство, и они служат причиной нормального ПДРФ. Выяснилось, что ПДРФ является четким генетическим признаком. Для ряда наследственных болезней описаны формы ПДРФ, прямо свидетельствующие о наличии заболевания или о носительстве патологически измененного гена. В настоящее время анализ ПДРФ стал одним из наиболее точных методов, используемых в генетике человека и медицинской генетике (< < Медицинская генетика> > ). Г. и. положила начало новому направлению исследований, получившему название «генетиканаоборот». Традиционный генетический анализ проводят в следующей последовательности: выбирается признак, устанавливается связь признака с генетической детерминантой и локализация этой детерминанты по отношению к уже известным. В «генетике наоборот» все происходит в обратном порядке. Из набора клонированных участков генома (или кДНК) с неизвестной функцией, характеризующихся определенным ПДРФ, выделяют те из них, которые наиболее тесно сцеплены с конкретным признаком. Если признаком является наследственное заболевание, с помощью выявления тесно сцепленного с ним полиморфного фрагмента можно осуществить диагностику этого заболевания по наличию той или иной формы ПДРФ. Этот подход позволил разработать методы ранней пренатальной диагностики, выявления носителей патологического гена в семьях даже для таких заболеваний, как хорея Гентингтона, болезнь Дюшенна, муковисцидоз, природа генетических дефектов при которых была абсолютно не известна. Затем можно точно определить положение соответствующего гена на хромосоме, выделить и проанализировать не только ген, но и продукты его так называемой экспрессии (мРНК и белок) в норме и при патологии. Полностью принципиальная схема «генетики наоборот» успешно реализована в случае болезни Дюшенна, муковисцидоза. На пути внедрения в медицинскую практику методов, используемых в Г. и., еще много трудностей технического порядка. Во многих лабораториях мира активно ведется разработка практически пригодных генно-инженерных диагностических методов, и можно надеяться, что такого рода методы уже в ближайшем будущем найдут применение для выборочного обследования групп повышенного риска в отношении наследственных болезней. Практическое значение Г. и. для медицины связано с перспективами исправления наследственных дефектов у человека, создания и использования микроорганизмов, потерявших свою патогенность, но сохранивших способность к формированию иммунитета. Разработаны методы синтеза антибиотиков, аминокислот, гормонов, витаминов, ферментов и т.д., основанные на использовании микроорганизмов, включивших соответствующие гены. Г. и. позволяет не только копировать природные соединения и процессы, но и модифицировать их, делать их более эффективными. Примером этого может служить новое направление исследований, названное белковой инженерией. Расчеты, производимые на основании данных об аминокислотной последовательности и пространственной организации молекул белков, показывают, что при определенных заменах некоторых аминокислотных остатков в молекулах ряда ферментов возможно значительное усиление их ферментативной активности. В изолированном гене, кодирующем синтез конкретного фермента, методами Г. и. проводят строго контролируемую замену определенных нуклеотидов. При синтезе ферментного белка под контролем такого модифицированного гена происходит заранее спланированная замена аминокислотных остатков в полипептидной цепи, что вызывает повышение ферментативной активности модифицированного фермента во много раз по сравнению с активностью природного прототипа. Из практических достижений Г. и. наиболее важными являются создание продуцентов биологически активных белков — инсулина, интерферона, гормона роста и др., а также разработка способов активизации звеньев обмена веществ, которые связаны с образованием низкомолекулярных биологически активных соединений. Таким путем получены продуценты ряда антибиотиков, аминокислот, витаминов, во много раз более эффективные, чем их продуценты, выведенные традиционными методами генетики и селекции.г и. разрабатываются способы получения чисто белковых вакцин против вирусов гепатита, гриппа, герпеса, ящура, реализована идея использования для вакцинации комбинированного вируса осповакцины, в геном которого встроены гены, кодирующие синтез белков других вирусов (например, вирусов гепатита или гриппа). В результате прививки таким вирусом организм получает возможность выработать иммунитет не только против оспы, но и против гепатита, гриппа или другого заболевания, вызываемого тем вирусом, синтез белка которого котируется встроенным геном. Как и любое достижение науки, успехи Г. и. могут быть использованы не только на благо, но и во вред человеку. Специально проведенные исследования показали, что опасность неконтролируемого распространения гибридных (рекомбинантных) ДНК не так велика, как представлялось ранее. Гибридные ДНК и несущие их бактерии оказались очень неустойчивыми к влияниям окружающей среды, нежизнеспособными в организме человека и животных при случайном проникновении. Известно, что в природе и без вмешательства человека имеются условия, которые обеспечивают обмен генетической информацией (так называемый поток генов). Однако на пути случайного проникновения в организм чужеродной генетической информации природа создала много эффективных барьеров. При работе с большинством гибридных молекул ДНК вполне достаточно обычных мер предосторожности, которые применяют, например, микробиологи при работе с инфекционным материалом. Для особых случаев разработаны эффективные способы биологической защиты и физической изоляции экспериментальных объектов от человека и окружающей среды. Библиогр.: Маниатис Г., Фрич Э. и Сэмбрук Дж. Молекулярное клонирование (методы генетической инженерии), пер. с англ., М., 1984; Уотсон Дж., Туз Дж. и Кури Ц. Рекомбинантные ДНК, пер. с англ., М., 1986.
|

 
 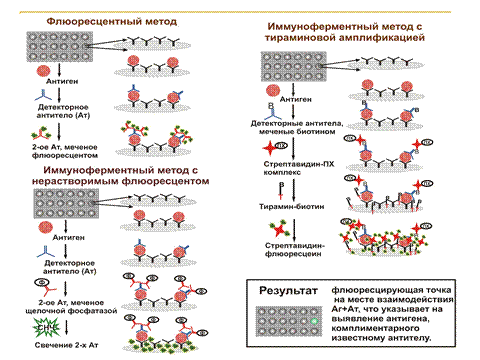
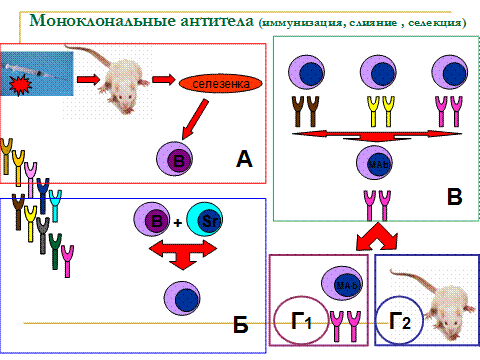

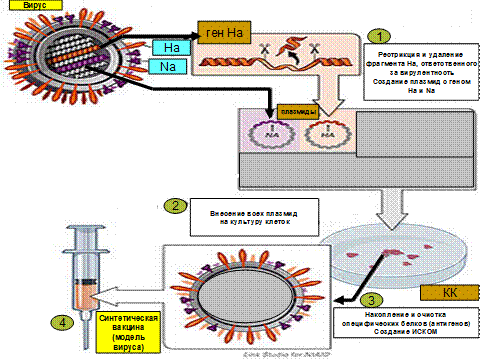
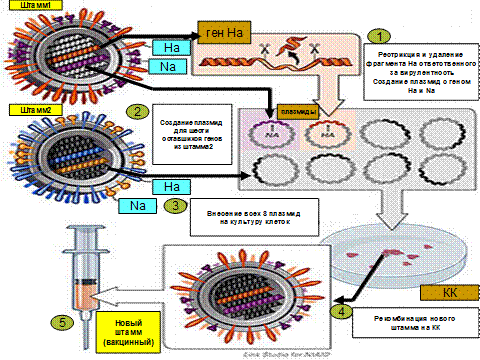
История генной инженерии
Генная инженерия появилась благодаря работам многих исследователей в разных отраслях биохимии и молекулярной генетики. На протяжении многих лет главным классом макромолекул считали белки. Существовало даже предположение, что гены имеют белковую природу. Лишь в 1944 году Эйвери, Мак Леод и Мак Карти показали, что носителем наследственной информации является ДНК. С этого времени начинается интенсивное изучение нуклеиновых кислот. Спустя десятилетие, в 1953 году Дж. Уотсон и Ф. Крик создали двуспиральную модель ДНК. Именно этот год принято считать годом рождения молекулярной биологии.
На рубеже 50 - 60-х годов были выяснены свойства генетического кода, а к концу 60-х годов его универсальность была подтверждена экспериментально. Шло интенсивное развитие молекулярной генетики, объектами которой стали E. coli, ее вирусы и плазмиды. Были разработаны методы выделения высокоочищенных препаратов неповрежденных молекул ДНК, плазмид и вирусов. ДНК вирусов и плазмид вводили в клетки в биологически активной форме, обеспечивая ее репликацию и экспрессию соответствующих генов. В 70-х годах был открыт ряд ферментов, катализирующих реакции превращения ДНК. Особая роль в развитии методов генной инженерии принадлежит рестриктазам и ДНК-лигазам.
Историю развития генетической инженерии можно условно разделить на три этапа.
Первый этап связан с доказательством принципиальной возможности получения рекомбинантных молекул ДНК invitro. Эти работы касаются получения гибридов между различнымиплазмидами. Была доказана возможность создания рекомбинантных молекул с использованием исходных молекул ДНК из различных видов и штаммов бактерий, их жизнеспособность, стабильность и функционирование.
Второй этап связан с началом работ по получению рекомбинантных молекул ДНК между хромосомными генами прокариот и различными плазмидами, доказательством их стабильности и жизнеспособности.
Третий этап - начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-рецепиента) генов эукариот, главным образом, животных.
Формально датой рождения генетической инженерии следует считать 1972 год, когда в Стенфордском университете П. Берг, С. Коэн, Х. Бойер с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli.
Читать дальше ► ферменты для манипуляций с ДНК и РНК
|
