|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Характеристика высокоэнергетических соединений
4.1 АТФ- универсальная форма химической энергии в клетке Слайд №28. У бактерий существуют три основных типа богатых энергией химических соединений: нуклеозидтрифосфаты, ацилфосфаты и ацилтиоэфиры (соединения с высокоэнергетической тиоэфирной связью, например, ацетилкознзим А, сукцинилкоэнзим А). Самую большую группу составляют соединения с высокоэнергетической фосфатной связью: ацилфосфаты, фосфорные эфиры енолов (фосфоенолпируват), нуклеотидди- и трифосфаты(аденозинтрифосфат, гуанозинтрифосфат (ГТФ), уридинтрифосфат (УТФ), цитидинтрифосфат (ЦТФ), дезокситимидинтрифосфат(дТТФ)), а также аденозинфосфосульфат, креатинфосфат.
Примеры некоторых таких богатых и бедных энергией соединений приведены в таблице 2. Эти соединения характеризуются тем, что одна из входящих в состав молекулы групп имеет высокий энергетический потенциал. При переносе этой группы происходит разрыв связи, соединяющей ее с молекулой, что приводит к резкому уменьшению свободной энергии, заключенной в молекуле химического соединения. Такие связи называются высокоэнергетическими, или макроэргическими. Присоединение группы с высоким энергетическим потенциалом к молекуле-акцептору повышает уровень ее свободной энергии, переводя молекулу в активированную форму, в которой это соединение может участвовать в биосинтетических реакциях. Долгое время считали, что единственный тип энергетической валюты — высокоэнергетические химические соединения, а среди них прежде всего АТФ, однако, оказалось, что клетка располагает тремя типами энергетической валюты: наряду с АТФ такую роль выполняют протонный и натриевый потенциалы на биологических мембранах. В то же время, неоспоримым является тот факт, что центральное место в процессах переноса химической энергии принадлежит системе аденозинтрифосфата. АТФ образуется в реакциях субстратного и мембранзависимого фосфорилирования. Слайд №29. Молекула АТФ содержит две макроэргические фосфатные связи, при гидролизе которых высвобождается значительное количество свободной энергии: АТФ + Н2 -> АДФ + Фн; AGo' = -31, 8 кДж/моль; АДФ + Н2 -> АМФ + Фн; AGo' = -31, 8 кДж/моль; Отщепление последней фосфатной группы от молекулы АМФ приводит к значительно меньшему высвобождению свободной энергии: АМФ + Н2 —> аденозин + Фн; AG0' = -14, 3 кДж/моль; Молекула АТФ обладает определенными свойствами, которые и привели к тому, что в процессе эволюции ей была отведена столь важная роль в энергетическом метаболизме клеток. Термодинамически молекула АТФ нестабильна, что вытекает из большой отрицательной величины свободной энергии (∆ G -свободной энергии) ее гидролиза. В то же время скорость неферментативного гидролиза АТФ в нормальных условиях очень мала, т.е. химически молекула АТФ высокостабильна. Последнее свойство обеспечивает эффективное сохранение энергии в молекуле АТФ, поскольку химическая стабильность молекулы препятствует тому, чтобы запасенная в ней энергия бесполезно рассеивалась в виде тепла. Малые размеры молекулы АТФ позволяют ей легко диффундировать в различные участки клетки, где необходим подвод энергии из вне для выполнения химической, осмотической или механической работы. И наконец, еще одно свойство молекулы АТФ, обеспечившее ей центральное место в энергетическом метаболизме клетки. Высвобождение свободной энергии при гидролизе АТФ составляет — 31, 8 кДж/моль. Если сравнить эту величину с аналогичными величинами для ряда других фосфорилированных соединений, то мы получим определенную шкалу. На одном из ее полюсов будут расположены фосфорилированные соединения, гидролиз которых приводит к высвобождению значительного количества свободной энергии (высокие отрицательные значения ∆ G- свободной энергии). Это так называемые " высокоэнергетические соединения". На другом полюсе будут располагаться фосфорилированные соединения, свободная энергия (∆ G) гидролиза которых имеет невысокое отрицательное значение (" низкоэнергетические" соединения). Пример высокоэнергетического соединения — фосфоенолпировиноградная кислота (∆ Go, = -58, 2 кДж/моль), низкоэнергетического — глицеро-1-фосфат (∆ Go'= —9, 2 кДж/моль). АТФ на этой шкале занимает промежуточное положение, что и дает ему возможность наилучшим образом выполнять энергетические функции: переносить энергию от высокоэнергетических к низкоэнергетическим соединениям.
4.2 Электрохимический трансмембранный градиент ионов (∆ μ н+) - вторая универсальная форма клеточной энергии
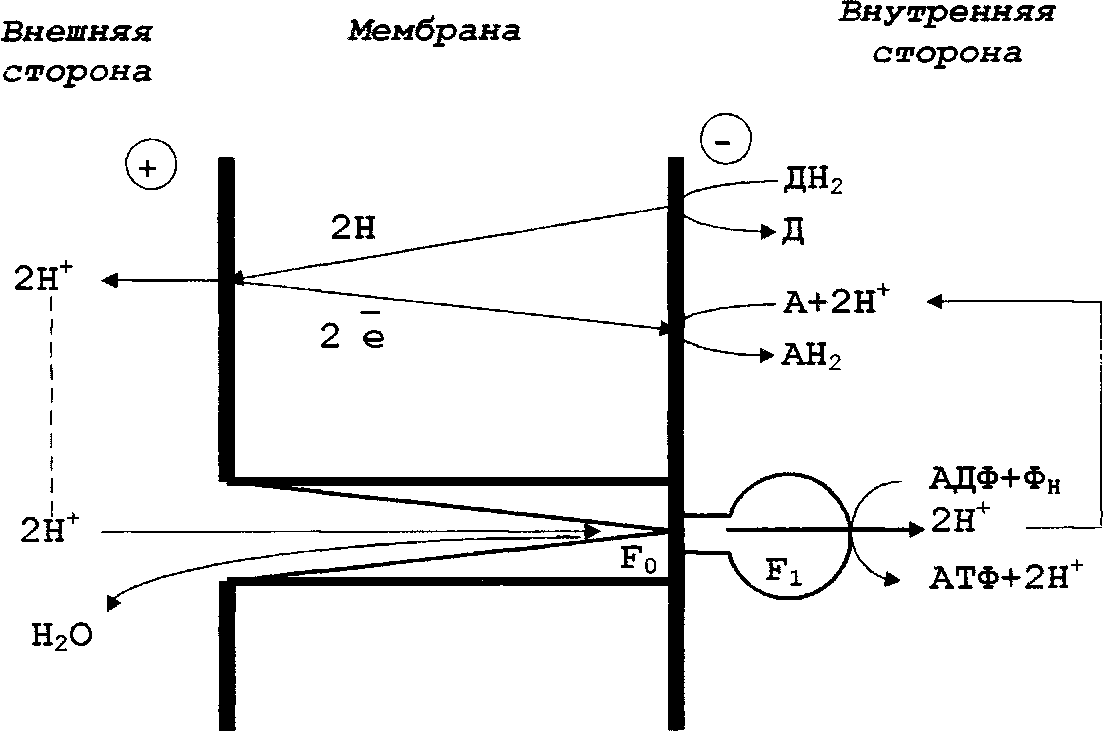
В течение длительного времени считали, что АТФ и другие высокоэнергетические соединения, находящиеся в равновесии с ним, представляют собой единственную форму энергии, которая может использоваться живыми клетками во всех энергозависимых процессах. Вопрос о характере связи между транспортом электронов, с одной стороны, и превращением фосфорных соединений, с другой, долгое время оставался неясным. Было установлено, что использование энергетических ресурсов (органических или неорганических соединений при дыхании, света - при фотосинтезе) связано с переносом электронов по цепи, состоящей из белковых и небелковых компонентов, способных к обратимому окислению и восстановлению. В результате этого переноса освобождающаяся на отдельных участках дыхательной или фотосинтетической цепи энергия трансформируется в химическую энергию фосфатных связей АТФ. Молекулярный механизм фосфорилирования, сопряженный с электронным транспортом, был неизвестен. Позднее были получены экспериментальные данные о существовании еще одной формы энергии, также используемой клеткой для совершения разного рода работы. Открытие этой формы энергии принадлежит английскому биохимику Питеру Митчеллу (P. Mitchell), разработавшему в 60-х гг. хемиосмотическую теорию энергетического сопряжения, объясняющую превращение(трансформацию) энергии, освобождающейся при электронном транспорте, в энергию фосфатной связи АТФ. Слайд №30. Согласно теории П. Митчелла при переносе электронов по окислительно-восстановительной цепи, локализованной в мембранах определенного типа, называемых энергопреобразующими, или сопрягающими, происходит неравномерное распределение протонов (Н+) в пространстве по обе стороны мембраны (рисунок 1).
Д - донор электронов, А - акцептор электронов, F0Fi - компоненты Н+-АТФ-синтазы Предложенная им модель предусматривает определенное расположение переносчиков электронов в сопрягающей мембране. В сопрягающих мембранах два атома водорода движутся от внутренней стороны цитоплазматической мембраны к наружной с помощью переносчика водорода (например, хинона). Затем два электрона возвращаются к внутренней стороне мембраны с помощью соответствующего электронного переносчика (например, цитохрома), а два протона освобождаются во внешнюю среду. Таким образом, в каждой окислительно-восстановительной цепи два атома водорода (Н+) переносятся из цитоплазмы клетки во внешнюю среду. Расположение переносчиков электронов в цитоплазматической мембране прокариот таково, что при работе любой электронтранспортной цепи (фотосинтетической или дыхательной) во внешней среде происходит накопление ионов водорода (протонов), приводящее к подкислению среды, а в клеточной цитоплазме — их уменьшение, сопровождающееся ее подщелачиванием, т.е. на мембране возникает ориентированный поперек (трансмембранный) градиент ионов водорода. Поскольку атомы водорода (H+)— химические частицы, несущие положительный заряд, неравномерное их накопление по обе стороны мембраны приводит к возникновению не только химического (концентрационного) градиента этих частиц, но и ориентированного поперек мембраны электрического поля (суммарный положительный заряд, где происходит накопление атомов Н+, и отрицательный заряд по другую сторону мембраны). Таким образом, при переносе электронов на цитоплазматической мембране возникает трансмембранный электрохимический градиент ионов водорода, обозначаемый символом (∆ μ н+ ), и измеряемый в вольтах (В, мВ), который состоит из электрического (трансмембранная разность электрических потенциалов ∆ ) и химического (концентрационного) компонентов (градиент концентраций Н+ — ∆ рН ). Итак, в соответствии с хемиосмотической теорией П. Митчелла, энергия, освобождаемая в результате работы электронтранспортной цепи, первоначально накапливается в форме трансмембранного градиента ионов водорода. Разрядка образующегося электрохимического градиента (∆ μ н+) происходит с участием локализованного в той же мембране протонного АТФ-синтазного комплекса: атомы Н+ возвращаются по электрохимическому градиенту ионов водорода ∆ μ н+ через Н+-АТФ-синтазу, при этом без возникновения каких-либо промежуточных высокоэнергетических соединений из АДФ и неорганического фосфата образуется АТФ. Слайд №31.Локализованная в мембране Н+-АТФ-синтаза катализирует реакции синтеза и гидролиза АТФ в соответствии с уравнением: АДФ + Фн (АТФ-синтаза) + пНнар+ (протоны) ↔ АТФ + Н20 + пНвнутр+ Реакция, протекающая слева направо, сопряжена с транспортом атомов Н+ по электрохимическому градиенту ионов водорода ( ∆ μ н+ ), что приводит к его разрядке и синтезу АТФ. Протекающая в противоположном направлении реакция гидролиза АТФ, сопровождающаяся переносом атомов водорода против градиента, приводит к образованию (или возрастанию) электрохимического градиента ∆ μ н+ на мембране. Таким образом, АТФ-синтазный ферментный комплекс служит механизмом, обеспечивающим взаимное превращение двух форм клеточной энергии: электрохимический градиент< -> АТФ (∆ μ н+ ↔ АТФ), устройством, сопрягающим процессы окислительной природы с фосфорилированием. Энергия в форме протонного градиента (∆ μ н+) может использоваться в различных знергозависимых процессах, локализованных на мембране. Синтез АТФ за счет протонного градиента (∆ μ н+) можно рассматривать как пример химической работы. С использованием энергии протонного градиента (∆ μ н+) могут осуществляться и другие виды химической работы в клетке: синтез пирофосфата, катализируемый связанным с мембраной ферментным комплексом; обратный перенос электронов, приводящий к восстановлению НАД(Ф)+ (никотинамидадинуклеотидафосфата). Энергия в форме протонного градиента (∆ μ н+) используется для поглощения ДНК в процессе генетической трансформации и для переноса белков через мембрану. Движение многих прокариот обеспечивается энергией протонного градиента (∆ μ н+). Важная роль принадлежит Слайд №32 Все это позволяет рассматривать энергию в форме протонного градиента (наряду с АТФ) как широко используемую внутри клетки. Преобразование энергии в клетке прокариот схематически изображено на рисунке 3. Как видно из этой схемы, АТФ и электрохимический градиент ионов водорода ∆ μ н+ можно считать двумя взаимно превращаемыми " энергетическими валютами" клетки, каждая из которых способна служить источником энергии для выполнения химической, осмотической, механической работ.
|
Последнее изменение этой страницы: 2017-04-12; Просмотров: 1745; Нарушение авторского права страницы