|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Физиологические свойства и особенности функционирования тканей
Основным свойством любой ткани является раздражимость, т.е. способность ткани менять свои физиологические свойства и функциональную активность в ответ на действие раздражителей. Раздражители – это факторы внешней или внутренней среды, действующие на живые ткани. Раздражители в зависимости от вида заключенной в них энергии могут быть механическими, химическими, электрическими и другими. К общим физиологическим свойствам тканей относятся возбудимость, проводимость, рефрактерность, лабильность. Возбудимость – способность живой ткани отвечать на действие раздражителя изменением физиологических свойств и возникновением процесса возбуждения. Возбудимыми тканями являются нервная, мышечная и секреторная (железистая). Мерой возбудимости является порог раздражения. Это минимальная сила раздражителя, которая вызывает видимые ответные реакции. Так как порог раздражения характеризует возбудимость, он может быть назван и порогом возбудимости. Раздражитель меньшей интенсивности, не вызывающий ответные реакции, называют подпороговым; раздражитель выше порогового значения – сверхпороговым. Проводимость – способность ткани передавать возникшее возбуждение за счет электрического сигнала, возникающего в ней. Рефрактерность – временное снижение возбудимости одновременно с возникшим в ткани возбуждением. Рефрактерность бывает абсолютной (нет ответа на любой раздражитель) и относительной (возбудимость постепенно восстанавливается, и ткань отвечает на сверхпороговый раздражитель). Лабильность – способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуются максимальным числом волн возбуждения, возникающих в ткани в единицу времени. Состояние активности и покоя возбудимых тканей. Возбудимая ткань может находиться в двух состояниях: покоя и активности. О состоянии покоя говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет видимого проявления ее функции. Состояние активности наблюдается в том случае, когда на ткань действует раздражитель, при этом изменяются уровень метаболизма и присущая ей функция. Состояние активности может проявляться в виде возбуждения и торможения. Возбуждение – это активный физиологический процесс, который возникает в ткани под действием раздражителя, при этом изменяются физиологические свойства ткани и активируются ее функции. При этом изменяются проницаемость клеточных мембран, соотношение ионных потоков, заряд клеточной мембраны, возникает потенциал действия, изменяется уровень метаболизма, повышается потребление кислорода и увеличивается выделение углекислого газа. Торможение – активный процесс, он возникает при действии раздражителей на возбудимую ткань и проявляется в подавлении другого возбуждения. Следовательно, функциональная активность ткани снижается или исчезает. Биоэлектрические явления возникают в тканях в процессе нормальной жизнедеятельности вследствие разности потенциалов между внутренней и наружной поверхностями клеточной мембраны. Согласно общепринятой мембранно-ионной теории причина возникновения разности потенциалов – неравномерное распределение ионов по обе стороны клеточной мембраны (в системе цитоплазма – окружающая среда). Клеточная мембрана состоит из двойного слоя фосфолипидов и слоя белков. Слой фосфолипидов прерывистый, белки клеточной мембраны подвижны и свободно плавают в фосфолипидном геле, они образуют каналы, через которые проходят различные вещества. В каналах есть специальный механизм, изменяющий свойства этих белков, – ворота канала, чтобы ионы могли проходить сквозь канал. Имеются каналы для калия, натрия, кальция, хлора. Они могут быть открыты или закрыты благодаря способности ворот открываться или закрываться под влиянием различных факторов. Это процесс управляемый. Клеточная мембрана характеризуется ионной асимметрией, т.е. разной концентрацией ионов снаружи и внутри клетки. Так, на внешней поверхности клеточной мембраны преобладают ионы натрия, на внутренней – калия.
Рис. 2.2. Итог работы насоса на одну молекулу АТФ
Ионная асимметрия поддерживается работой натрий-калиевого насоса – механизма, обеспечивающего активный транспорт ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na+ внутри клетки и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами К+, находящимися вне клетки, и переносит их в цитоплазму (рис. 2.2). Энергия, необходимая для транспорта ионов, освобождается при расщеплении АТФ. Работа натрий-калиевого насоса обеспечивает внутри клетки высокую концентрацию ионов калия и низкую концентрацию ионов натрия. Возникновение потенциала – результат избирательной проницаемости мембраны. В состоянии покоя клеточная мембрана хорошо проницаема для ионов калия, хлора и почти непроницаема для ионов натрия. Мембранный потенциал, или потенциал покоя. В состоянии покоя ионы калия выходят из клетки на наружную поверхность клеточной мембраны по градиенту концентрации (диффузия ионов) – пассивный транспорт через открытые калиевые каналы. Под влиянием ионов калия на наружной поверхности увеличивается положительный заряд. Ионы натрия почти не проникают внутрь клетки, так как каналы для них в состоянии покоя закрыты. Анионы органических соединений из-за больших размеров не проникают через клеточную мембрану, остаются внутри клетки, создавая там отрицательный заряд. Таким образом, в состоянии покоя снаружи создается положительный заряд, а внутри – отрицательный. Этот заряд называется мембранным потенциалом покоя. Он равен 60–90 м В. Мембранный потенциал обеспечивает состояние «боевой готовности» клетки для ее реакции на действие раздражителей. Ионные механизмы возникновения потенциала действия. Потенциал действия – это сдвиг потенциала, возникающий на мембране клетки при действии порогового и сверхпорогового раздражителей, что сопровождается перезарядкой мембраны, в результате из состояния покоя клетка переходит в состояние активности. При действии порогового или сверхпорогового раздражителя прежде всего открываются натриевые, а затем и дополнительные калиевые каналы. Следствием является увеличение проницаемости клеточной мембраны для этих ионов. Для ионов натрия она повышается в 400–500 раз, для ионов калия – в 10–15 раз. Через открывшиеся каналы ионы натрия пассивно движутся внутрь клетки по электрохимическому градиенту, что приводит к деполяризации мембраны, а ионы калия диффундируют на наружную ее поверхность. В итоге развиваются следующие процессы: 1) локальный ответ (местное возбуждение); 2) высоковольтный пиковый потенциал (спайк); 3) следовые колебания. Локальный ответ может быть самостоятельной формой ответа ткани, если раздражитель не достигает порогового значения, или начальной частью потенциала действия, если раздражитель равен порогу раздражения или превышает его. Потенциал действия включает несколько фаз. Локальный ответ – это незначительное снижение потенциала мембраны, которое исчезает, как только раздражитель перестает действовать. Если же действие раздражителя достаточно сильное и длительное, то отрицательный заряд внутри клетки постепенно снижается (за счет поступления в цитоплазму положительно заряженных ионов натрия) и затем меняется на положительный. Снаружи клетки остаются отрицательно заряженные анионы, которые не могут пройти через ее мембрану вслед за натрием, поэтому положительный заряд внешней поверхности уменьшается до нуля и сменяется на противоположный – фаза деполяризации. После достижения на внутренней поверхности мембраны заряда, равного 20–30 мв, каналы для натрия закрываются, активизируется работа калий-натриевого насоса и заряд клеточной мембраны восстанавливается – фаза реполяризации. Изменение заряда клеточной мембраны во время ее возбуждения показано на рис. 2.3.
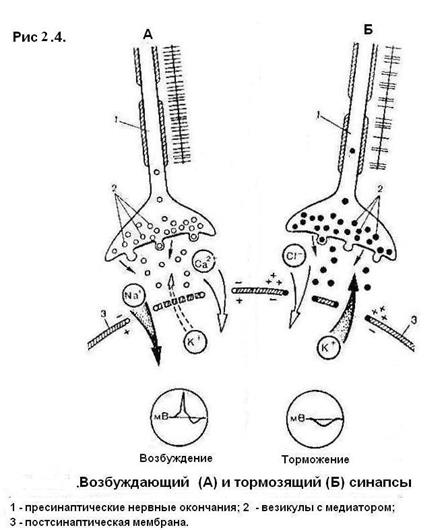
Рис 2.3. Потенциал действия нервной клетки: 1 – …5 ? Синапс Синапс – это структурно-функциональное образование, обеспечивающее переход возбуждения с окончания нервного волокна на иннервируемую клетку (рис. 2.4). Синапсы различают по способу передачи импульса (электрические, химические, смешанные), по локализации (центральные, периферические), по виду процессов на постсинаптической мембране (возбуждающие, тормозящие).
Рис. 2.4. Структура синапса: 1 – пресинаптическое нервное окончание; 2 – нейротрубочки; 3 – митохондрии; 4 – везикулы с медиатором; 5 – пресинаптическая мембрана; 6 – синаптическая щель; 7 – постсинаптическая мембрана; 8 – эффекторная клетка
Рассмотрим работу нервно-мышечного синапса. Нервный импульс по аксону направляется к иннервируемой структуре, достигает конечного участка (терминали) аксона и деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы и ионы кальция из среды, окружающей синапс, входят внутрь терминали аксона. Движение везикул (пузырьков, содержащих медиатор) упорядочивается по направлению к пресинаптической мембране. Ионы кальция стимулируют движение везикул. Достигая пресинаптической мембраны, везикулы разрываются – освобождается ацетилхолин, который поступает в синаптическую щель и взаимодействует с рецепторами на постсинаптической мембране. В результате открываются натриевые каналы и формируется потенциал концевой пластины. Медиаторы – это химические вещества, которые участвуют в передаче возбуждения или торможения в химических синапсах с пресинаптической мембраны на постсинаптическую мембрану. Важная особенность рецепторов постсинаптической мембраны – их способность вступать в биохимическое взаимодействие только с определенным видом медиатора. Межнейронные синапсы мало отличаются от нервно-мышечных. Каналы межнейронных синапсов в мозге регулируются различными медиаторами, поэтому в них могут развиваться как возбуждающие, так и тормозящие процессы. На постсинаптической мембране возбуждающих синапсов под действием медиатора происходит местная деполяризация, которая, суммируясь с аналогичным процессом на других постсинаптических мембранах этого нейрона, может привести к развитию потенциала действия на аксоне нейрона. На постсинаптической мембране тормозящих синапсов развивается процесс гиперполяризации, т. е. увеличения разности потенциалов, что приводит к снижению возбудимости нейрона. В отличие от мышечной клетки скелетной мускулатуры, которая образует одно нервно-мышечное соединение с аксоном нервной клетки, к телу каждого из нейронов, входящих в состав нервного центра, подходит не большое количество нервных окончаний от многочисленных нервных клеток. И в результате на теле каждого нейрона образуется большое число (до 10 тысяч) так называемых аксодендритных и аксосоматических синапсов. Одни из них служат для проведения возбуждения через нервный центр и называются возбуждающими синапсами, а другие – для блокады проведения возбуждения через этот центр и называются тормозящими. В возбуждающих контактах возбуждение передается с помощью медиаторов, которые вызывают местную деполяризацию на постсинаптической мембране. Но для возбуждения нейрона, воспринимающего информацию, недостаточно вызвать деполяризацию в одном контакте. Только при одновременном возбуждении многих участков на постсинаптической мембране деполяризация может достигнуть нужной величины и возбуждение сможет распространяться далее по аксону к следующему нейрону. Тормозящие контакты служат для блокирования проведения возбуждения через нервный центр. Специальные нейроны выделяют медиатор, который на постсинаптической мембране того же воспринимающего нейрона вызывает не деполяризацию, а гиперполяризацию мембраны. Тормозящим медиатором в ЦНС может быть аминокислота глицин, гамма-аминомасляная кислота и др. Под влиянием этих веществ на постсинаптической мембране открываются хлорные каналы, анионы хлора по электрохимическому градиенту поступают в клетку, увеличивая на ней разность потенциалов. В результате возбудимость нейрона понижается и он не может более воспринимать и передавать далее возбуждающий сигнал. Такое торможение называется постсинаптическим гиперполяризационным. Торможение в ЦНС было открыто И.М. Сеченовым (1863 г.), который экспериментально установил наличие в зрительных буграх головного мозга лягушки нервных механизмов, которые угнетают спинальные рефлексы. Значение торможения для жизнедеятельности организма состоит в том, что оно блокирует беспрепятственное распространение возбуждения на многочисленные нервные центры, морфологически связанные друг с другом, и направляет его по определенным нервным путям, осуществляя регулирующую, координирующую функцию, в результате которой в первую очередь осуществляются биологически наиболее важные рефлекторные реакции, а менее важные тормозятся.
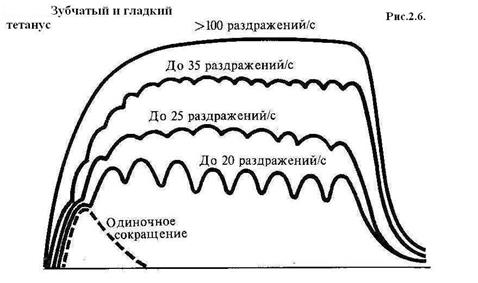
Рис. 2.5. Строение возбуждающего (А) и тормозящего (Б) синапсов: 1 – пресинаптические нервные окончания; 2 – везикулы с медиатором; 3 – постсинаптическая мембрана Скелетные мышцы Основу скелетных мышц формируют тонкие пучки поперечно-полосатых мышечных волокон, образующих мясистое брюшко, которое может активно сокращаться. На обоих концах оно переходит в сухожилие и прикрепляется к костям. Каждое мышечное волокно (миофибрилла) представляет собой многоядерную клетку, в состав которой входят мышечные волоконца (протофибриллы), или сократительные мышечные белки, упорядоченное расположение которых и придает мышце поперечную исчерченность. Виды мышечных сокращений: – изотоническое (изменяется длина волокон при неизменном напряжении; – изометрическое (изменяется напряжение при неизменной длине волокон); – смешанное (изменяется и напряжение и длина мышечных волокон). В организме в естественных условиях чаще всего происходит смешанное сокращение. Мышца может сокращаться в одиночном режиме, когда на нее действуют одиночные раздражители, или в режиме непрерывного длительного укорочения – тетануса, когда раздражители действуют с меньшим интервалом, чем длится одно полное сокращение. Тетанус может быть гладким и зубчатым. Гладкий возникает при большой частоте раздражения, при которой мышца не успевает даже частично расслабиться перед следующим сокращением. Зубчатый тетанус развивается при более редкой частоте раздражений, когда мышца успевает частично расслабиться к моменту следующего раздражения (рис. 2.6 и 2.7).
Рис. 2.6. Одиночное мышечное сокращение: 1 – точка раздражения; 1–2 – латентный период; 2–3 – фаза сокращения; 3–4 – фаза расслабления
Рис. 2.7. Зубчатый и гладкий тетанусы
Механизм мышечного сокращения и расслабления. Мышечное волокно содержит два типа белков: сократительные и модуляторные (регуляторные). К сократительным относятся актин и миозин, расположенные в виде толстых и тонких нитей параллельно друг другу. Скольжение актиновых нитей вдоль миозиновых и обеспечивает укорочение мышцы, т.е. ее сокращение (рис. 2.8). В состоянии покоя такого скольжения не происходит, так как взаимодействие сократительных белков блокируется тропонином и тропомиозином (модуляторными белками). При действии раздражителя модуляторные белки меняют свою конфигурацию так, что перестают блокировать сократительные белки и не препятствуют соединению активных центров на нитях актина с выростами (мостиками) на нитях миозина и последующему скольжению нитей актина. Изменение конфигурации тропонина и пропомиозина возможно только в присутствии ионов кальция. Хранилищем (депо) ионов кальция в мышце являются цистерны саркоплазматического ретикулюма (эндоплазматической сети в мышце). В мембранах этих цистерн при развитии в них потенциала действия открываются кальциевые каналы и освободившиеся ионы взаимодействуют с модуляторными белками, создавая условия для скольжения нитей актина. Энергию для этого скольжения дает распад АТФ, который происходит при взаимодействии миозиновых мостиков с активными центрами. Расслабление мышцы также является активным процессом и требует затраты энергии АТФ. Энергия расходуется на работу кальциевого насоса, который против концентрационного градиента возвращает ионы кальция в саркоплазматический ретикулюм. Это приводит к инактивации активных центров на нитях актина и отделению их от миозиновых мостиков. Доказательством активности процесса расслабления служит трупное окоченение, т. е. развитие мышечной контрактуры после смерти, когда отсутствие энергии для работы насосов не позволяет мышцам расслабляться. Эти же механизмы вызывают контрактуру при утомлении, так как при длительной работе слишком много энергии тратится на мышечное сокращение и ее не хватает для возвращения ионов кальция в саркоплазматический ретикулюм. На рис. 2.8 показано строение поперечно-полосатого мышечного волокна и основные механизмы ее сокращения. Скелетные мышцы состоят из поперечно-полосатой скелетной мускулатуры. Основной единицей мышечной ткани является мышечное волокно, которое, в свою очередь, состоит из протофибрилл – пучков сократимых белков (актина и миозина), способных вдвигаться друг в друга. Сокращение мышцы (вхождение актина и миозина друг в друга) происходит только при наличии АТФ и ищнов кальция. Запускается этот процесс в мышце под действием нервных импульсов, поступающих от нервной системы.
Рис. 2.8. Строение мышечной ткани: 1 – мышечное волокно; 2 – пучки сократительных белков; 3 – актин; 4 – миозин
Нервная система Структурная единица нервной ткани – нейрон – имеет тело и отростки. По коротким (обычно многочисленным) отросткам – дендритам – возбуждение передается телу нервной клетки, а по длинному одиночному аксону оно распространяется к другим нейронам или рабочим органам (мышцам и железам). Передача возбуждения с одной клетки на другую осуществляется при помощи синапсов.
Рис. 2.9. Строение нейрона: 1 – аксонный холмик; 2 – тело; 3 – пресинаптические окончания; 4 – дендриты; 5 – аксон; 6 – перехват Ренвье; 7 – миелиновая оболочка
Центральная нервная система . Физиология нервных центров. Центральная нервная система состоит из головного и спинного мозга. Основные отделы головного мозга представлены на рис. 2.10.
Рис. 2.10. Строение головного мозга: 1 – верхний участок спинного мозга; 2 – продолговатый мозг; 3 – мост; 4 – мозжечок; 5 – средний мозг; 6 – четверохолмие; 7 – промежуточный мозг; 8 – кора больших полушарий; 9 – мозолистое тело, соединяющее правое полушарие с левым; 10 – перекрест зрительных нервов; 11 – обонятельные луковицы
Нервным центром называется совокупность нервных клеток, расположенных в определенном отделе центральной нервной системы и регулирующих определенную функцию. Нервные центры в зависимости от расположения подразделяются на спинальные (в спинном мозге), бульбарные (в продолговатом мозге), мезенцефальные (в среднем мозге) и др. Нервные центры подразделяются также на соматические (регулирующие сокращения скелетной мускулатуры) и вегетативные (регулирующие работу внутренних органов). В нервном центре все нейроны связаны между собой с помощью синапсов, поэтому их свойства в значительной степени определяются свойствами последних. Для них характерны одностороннее проведение возбуждения, синаптическая задержка проведения возбуждения, низкая лабильность и высокая утомляемость. Как и в синапсах, в нервных центрах имеет место суммация возбуждения, т. е. сложение местных ответов при последовательном или одновременном поступлении нескольких импульсов к одному нейрону. За счет синаптических связей импульс, поступивший к одному нейрону, может распространяться на другие, рядом лежащие, что способствует распространению возбуждения. Такое явление называется иррадиацией. Противоположное явление, т. е. схождение импульсов от нескольких нейронов к одному, называется конвергенцией. Конвергенция лежит в основе суммации. В нервных центрах может развиваться не только возбуждение, но и торможение. Усиление активности органа или отдела ЦНС называется возбуждением, а снижение (когда нейрон не способен к выработке нервных импульсов) – торможением. Торможение – активный процесс, возникающий при действии раздражителей на нервные центры и проявляющийся в подавлении другого возбуждения. В результате функциональная активность центра снижается или исчезает. К физиологическим свойствам нервных волокон относятся: · возбудимость – способность приходить в состояние возбуждения в ответ на раздражение; · проводимость – способность передавать нервные возбуждения в виде потенциала действия от места раздражения по всей длине; · рефрактерность – способность временно резко снижать возбудимость в процессе возбуждения.
Главная функция нервных волокон – проведение нервных импульсов. По особенностям строения и функциям нервные волокна подразделяются на два вида. Так, безмиелиновые нервные волокна состоят из осевого цилиндра, покрытого оболочкой. Их диаметр 5–7 мкм, скорость проведения импульса 1 – 2 м/с. Миелиновые волокна состоят из осевого цилиндра, покрытого кроме обычной оболочки еще и миелиновой оболочкой. Осевой цилиндр имеет мембрану и аксоплазму. Миелиновая оболочка на 80 % состоит из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье). При диаметре 12–20 мкм скорость проведения возбуждения составляет 70–120 м/с. Механизм проведения возбуждения. В безмиелиновых нервных волокнах (рис. 2.11) возбуждение распространяется за счет малых круговых ионных токов, возникающих между возбужденным и невозбужденным участками волокна. Возбужденный участок мембраны на наружной поверхности заряжается отрицательно, а невозбужднный снаружи заряжен положительно. Ток передается от положительного участка к отрицательному. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона. Такой путь передачи возбуждения энергетически невыгоден, так как требует большой затраты энергии для поддержания ионных градиентов на большой площади. В миэлиновых волокнах при раздражении возбуждение возникает только в области перехвата Ранвье, где и развивается деполяризация. Соседний перехват в это время поляризован, и между ними возникает разность потенциалов, приводящая к развитию круговых токов. При этом возбуждение распространяется скачкообразно, от одного перехвата к другому. Такой способ распространения более экономичен, требует меньших затрат энергии калий-натриевого насоса, поэтому скорость проведения в миэлиновых волокнах значительно выше.
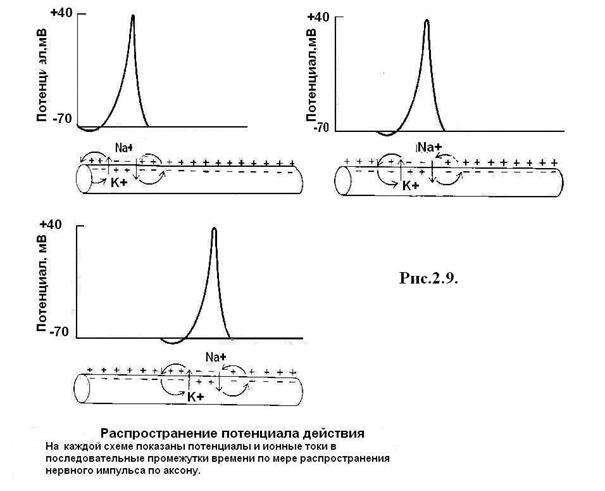
Рис. 2.11. Проведение возбуждения по нервному волокну. На каждой схеме показаны потенциалы действия и ионные токи по мере распространения нервного импульса по аксону
Законы проведения возбуждения по нервному волокну: 1. Закон анатомо-физиологической целостности. Проведение возбуждения по нервному волокну возможно лишь в случае, если не нарушена его целостность и сохранены его физиологические свойства. Сдавление, охлаждение, использование анестетиков делают проведение по нерву невозможным. 2. Закон изолированного проведения возбуждения. Возбуждение распространяется только вдоль нервного волокна, не распространяясь на соседние, которые находятся в этом же нервном стволе. 3. Закон двустороннего проведения возбуждения. Возбуждение распространяется вдоль нервного волокна в обе стороны от места приложения раздражителя. Нервная система человека Единая нервная система условно подразделяется на две части: соматическую (обеспечивает управление скелетной мускулатурой) и вегетативную. Вегетативная нервная система. Это часть нервной системы, которая обеспечивает управление работой всех внутренних органов и систем, желез внутренней секреции, кровеносных и лимфатических сосудов и других органов (см. таблицу). Она также координирует деятельность всех внутренних органов, регулирует обменные, трофические процессы во всех органах и частях тела человека, поддерживает постоянство внутренней среды. По своей функции вегетативная нервная система неподконтрольна нашему сознанию, но находится в подчинении ЦНС. |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 469; Нарушение авторского права страницы