|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Газообмен в легких и тканях
В легких происходит обмен газов между воздухом, находящимся в альвеолах, и притекающей к легким по сосудам малого круга венозной кровью. При спокойном дыхании объем альвеолярного воздух соответствует ФОЕ. Если в среднем остаточный объем легких равен 1200 мл, а резервный объем выдох выдоха – 1300 мл, то в легких при спокойном вдохе и выдохе содержится 1200 + 1300 = 2500 мл воздуха. Этот воздух заполняет альвеолы и нижние дыхательные пути и составляет газовую среду организма. Каждый раз при спокойном вдохе в легкие поступает 500 мл воздуха. Из них часть заполняет анатомическое мертвое пространство (около 175 мл). Оно называется так потому, что воздух, находящийся в нем, заполняет носоглотку, трахею и бронхи; он не доходит до альвеол и не обменивается с кровью. Следовательно, до альвеол доходит около 325 мл воздуха, и всякий раз ФОЕ обновляется примерно на 1/7 часть. Этот показатель называется коэффициентом легочной вентиляции (КЛВ). Воздух по дыхательным путям поступает в альвеолы – воздушные ячейки, густо оплетенные сетью кровеносных капилляров, несущих венозную кровь; она содержит мало кислорода и относительно много углекислого газа. Газообмен в легких представляет собой процесс перехода кислорода из легочных альвеол в венозную кровь, а углекислого газа – из крови в альвеолы. В результате венозная кровь, поступившая к легким по легочной артерии, превращается в артериальную и по легочным венам направляется к сердцу. Газообмен происходит путем физической диффузии газов. Движущей силой диффузии является разность концентраций. В данном случае это будет разность парциальных давлений кислорода и углекислого газа по обе стороны аэрогематического барьера: по градиенту парциальных давлений кислород из альвеолярного воздуха переходит в венозную кровь, а углекислый газ диффундирует из крови в альвеолы.
Таблица 4.1 Газовый состав атмосферного, альвеолярного и выдыхаемого воздуха
Примечание. Содержание дано в процентах, парциальное давление (в скобках) – в мм рт. ст. Парциальное давление – это давление газа, находящегося в смеси с другими газами, пропорциональное его процентному содержанию. Градиент парциального давления кислорода и углекислого газа – это сила, с которой молекулы этих газов стремятся проникнуть через альвеолярно-капиллярную мембрану в кровь. Давление газа, растворенного в жидкости, обозначают как напряжение.
Рис. 4.5. Газообмен между альвеолами и легочными капиллярами
Парциальное давление кислорода в альвеолярном воздухе (при его процентном содержании 14,2 %) равно примерно 100 мм рт. ст., а углекислого газа (при его процентном содержании 5,5 %) – 40 мм рт. ст. Напряжение кислорода в венозной крови составляет 40 мм рт. ст., а углекислого газа – 46 мм рт. ст. Кислород по градиенту парциального давления следует из области более высокого давления (из альвеол) в область низкого и попадает венозную кровь, а углекислый газ переходит из венозной крови в альвеолы. В результате от легких оттекает артериальная кровь, насыщенная кислородом и лишенная углекислого газа (рис. 4.5). Вентиляция легких все время обновляет состав альвеолярного воздуха примерно на 1/7. Благодаря вентиляции процентное содержание кислорода и углекислого газа и, стало быть, парциальное давление этих газов, поддерживается на постоянном уровне, что, в свою очередь, обеспечивает постоянство газового состава крови, а непрерывный кровоток и диффузия газов через альвеолярно-капиллярную мембрану позволяют постоянно превращать венозную кровь в артериальную (табл. 4.1).
Транспорт газов кровью Кислород от легких к тканям и углекислый газ от тканей к легких переносится кровью. В основном эти газы находятся в крови в химически связанном состоянии. Только небольшая их часть может находиться в свободном состоянии, формируя напряжение кислорода и углекислого газа в крови. В легких кислород переходит из альвеол сначала в плазму крови легочных капилляров, а затем поступает в эритроциты, где образует с гемоглобином эритроцитов обратимое соединение – оксигемоглобин: Нв + О2 = НвО2. Данная реакция является обратимой, а равновесие может сдвигаться вправо или влево в зависимости от имеющейся концентрации кислорода и ряда других факторов. Максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови (КЕК). Кислородная емкость крови зависит от содержания в ней гемоглобина. Каждый грамм гемоглобина способен присоединить 1,34 мл кислорода. При содержании в крови 140 г/л гемоглобина кислородная емкость крови будет 1,34 ·140 = около 190–200 мл кислорода.. Так как в тканях кислород непрерывно расходуется, его концентрация (парциальное напряжение) там ниже, чем в артериальной крови. В результате происходит распад (диссоциация) оксигемоглобина на гемоглобин и кислород. Кислород растворяется в плазме и поступает в тканевую жидкость к клеткам тканей, а гемоглобин связывается с поступившим из тканей углекислым газом, образуя также обратимое соединение карбгемоглобин. В виде карбгемоглобина транспортируется лишь 5–15 % всего углекислого газа. Основная часть углекислого газа переносится кровью в форме бикарбонатов натрия и калия (80–90 %), причем бикарбонат калия образуется в эритроцитах, а бикарбонат натрия – в плазме крови. В легких происходит распад бикарбонатов и карбгемоглобина и образовавшийся углекислый газ поступает в альвеолы и выводится из организма в окружающую среду. Одновременно из альвеол в кровь поступает кислород, который связывается с гемоглобином. Эти отношения могут быть выражены протекающими в эритроцитах обратимыми химическими реакциями: Н2О + СО2 = Н2СО3 (реакцию ускоряет в 20 000 раз фермент карбоангидраза), КНвО2 + Н2СО3 = КНв +О2 + КНСО3-.
Газообмен в тканях. Артериальная кровь с напряжением кислорода 100 мм рт. ст. и углекислого газа 40 мм рт. ст. направляется от легких в ткани, где в результате непрерывно происходящих процессов биологического окисления используется кислород и образуется углекислый газ. В клетках тканей напряжение кислорода колеблется от 0 до 20 мм рт. ст., а напряжение углекислого газа в тканевой жидкости составляет около 60 мм рт. ст. За счет разности напряжений углекислый газ диффундирует из тканевой жидкости в кровь, а кислород переходит из крови в ткани. В результате артериальная кровь превращается в венозную и по венам поступает в легкие, где цикл обмена газов повторяется (рис. 4.6).
Рис. 4.6. Газообмен между кровью и тканями Регуляция дыхания Регуляция дыхания представляет собой процесс управления первым этапом дыхания – легочной вентиляцией путем изменения глубины и частоты дыхания. Эти параметры могут регулироваться автоматически и произвольно. Фундаментом регуляции является автоматическая регуляция дыхания. Доказательством этого служит тот факт, что ритмические дыхательные движения совершаются и во сне и при бодрствовании, не требуя участия нашего сознания. Однако человек может до определенного предела произвольно изменять характер дыхания. Автоматическая регуляция дыхания в любых условиях жизнедеятельности обеспечивает состояние «эйпноэ», т. е. «хорошего» комфортного дыхания, интенсивность которого точно соответствует метаболическим потребностям организма в кислороде и выведении углекислого газа. Регуляцию дыхания обеспечивает дыхательный центр, который генерирует (задает) дыхательный ритм, т. е организует сокращение дыхательных мышц с определенной силой и частотой так, чтобы вдох плавно переходил в выдох и чтобы частота и глубина дыханий в покое и в меняющихся условиях жизнедеятельности обеспечивали метаболические потребности организма. А это, в свою очередь, возможно при поддержании постоянства газового состава крови.
Рис. 4.7. Эфферентные связи дыхательного центра продолговатого мозга.
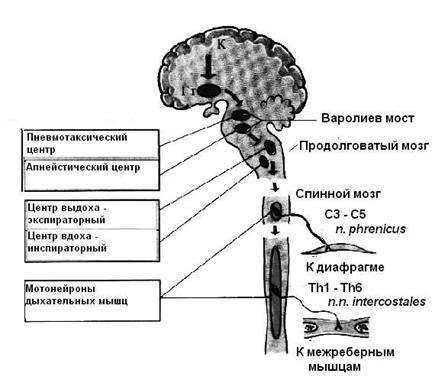
Дыхательный центр представляет собой совокупность нейронов, которые объединены общей функцией и расположены на разных «этажах» ЦНС (рис. 4.7). Выделяют четыре «этажа»: спинной мозг, продолговатый мозг, варолиев мост, кора больших полушарий. Каждый из перечисленных отделов выполняет определенную функцию. Спинальный уровень представлен просто двигательными нейронами передних рогов спинного мозга, аксоны которых образуют соматические нервы, иннервирующие дыхательные мышцы. Это диафрагмальный и межреберные нервы. Этот компонент не имеет самостоятельного значения, так как подчиняется импульсам из вышележащих отделов. Дыхательный центр продолговатого мозга обеспечивает последовательную смену вдоха и выдоха. Он представляет собой сложное образование, состоящее из двух отделов – центра вдоха (инспираторный отдел) и центра выдоха (экспираторный отдел). Нейроны продолговатого мозга могут самопроизвольно генерировать нервные импульсы (обладают автоматизмом). В верхней части варолиевого моста находится так называемый пневмотаксический центр, который контролирует деятельность расположенных ниже дыхательных центров вдоха и выдоха и обеспечивает нормальные дыхательные движения В процессах регуляции дыхания принимают участие также вышележащие отделы ЦНС, которые обеспечивают тонкие приспособительные изменения дыхания при различных видах деятельности организма. Важная роль в регуляции дыхания принадлежит большим полушариям головного мозга и их коре, благодаря которой произвольно осуществляется приспособление дыхательных движений при разговоре, пении, спорте, трудовой деятельности человека. Таким образом, в регуляции дыхательного процесса принимают участие различные структуры мозга, обеспечивая участие дыхательной системы в поведенческих реакциях, изменение дыхания при эмоциях, участие дыхания при речи, пении и т. д., но ведущую роль в регуляции легочной вентиляции играет дыхательный центр продолговатого мозга (рис. 4.8).
Рис. 4.8. Структура дыхательного центра Основную роль в регуляции смены вдоха и выдоха и в регуляции глубины дыхания играют потоки импульсов, поступающие в дыхательный центр от хемо- и механорецепторов. Основным регулятором активности ДЦ является информация о газовом составе крови и цереброспинальной жидкости (ликвора), которая поступает соответственно от периферических (артериальных) и центральных (мозговых) хеморецепторов. Периферические хеморецепторы находятся в каротидном и аортальном тельцах, а центральные – в продолговатом мозге, неподалеку от самого ДЦ. Они реагируют на повышение напряжения углекислого газа (СО2) и повышение концентрации водородных ионов, на понижение напряжения кислорода, но главным стимулом, управляющим дыханием, является содержание СО2 в крови и во внеклеточной жидкости мозга. Центральные хеморецепторы очень чувствительны к изменениям рН и напряжения СО2. Импульсы от нейронов дыхательного центра передаются к мотонейронам спинного мозга, управляющим наружными межреберными мышцами и диафрагмой (рис. 4.9).
Рис. 4.9. Механизм вдоха После вдоха автоматически происходит выдох. Для развития выдоха необходимо снижение активности центра вдоха и активация центра выдоха. Это происходит потому, что растяжение легочных альвеол во время вдоха вызывает раздражение рецепторов. В этом принимают участие несколько групп механорецепторов, особенно большая роль принадлежит рецепторам растяжения легких, реагирующим на растяжение стенок конечных отделов воздухоносных путей при вдохе. Возбуждение этих рецепторов возрастает при увеличении объема легких при вдохе, импульсы от них по ветвям блуждающих нервов направляются центру выдоха (экспираторным нейронам) и вызывают инспираторно-тормозящий рефлекс: вдох обрывается, и происходит выдох. Это известный рефлекс Геринга–Брейера: вдох порождает выдох. При двусторонней перерезке блуждающих нервов не происходит своевременной смены фаз дыхания: вдох становится более продолжительным (рис. 4.10).
Рис. 4.10. Изменение дыхания после двусторонней ваготомии
Схема нейронной организации дыхательного цикла представлена на рис. 4.11.
Рис. 4.11. Структура дыхательного цикла
Инспираторные нейроны получают возбуждающий импульс от центральных и периферических хеморецепторов и передают команду мотонейронам спинного мозга и через них – мышцам вдоха: происходит активный вдох. Одновременно активируются экспираторные нейроны, которые через тормозные нейроны затормаживают инспираторные нейроны, срабатывает механизм выключения вдоха, и пассивно происходит выдох. Дыхательный центр не только обеспечивает ритмическое чередование вдоха и выдоха, но и способен изменять глубину и частоту дыхательных движений, приспосабливая тем самым легочную вентиляцию к текущим потребностям организма. В осуществлении этой функции важная роль принадлежит пневмотаксическому центру, расположенному в варолиевом мосту. Активность дыхательного центра зависит от потребности организма в кислороде. В покое она невелика и увеличивается при физической работе или эмоциональном возбуждении. Чем больше в организме накапливается углекислого газа, тем активней он выводится. Таким образом, в регуляции дыхания участвуют различные как нервные, так и гуморальные механизмы, которые автоматически подбирают глубину и частоту дыхания, создавая оптимальные условия для газообмена в любых условиях. Глава 5. Пищеварение |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 401; Нарушение авторского права страницы