|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
МЕТАБОЛИЧЕСКОЕ НАПРАВЛЕНИЕ ЭВОЛЮЦИИ МИКРООРГАНИЗМОВ
Пробионты, появившиеся 3, 6 млрд лет назад, были первичными гетеротрофами и получали энергию при расщеплении органических веществ абиогенного происхождения, в изобилии имевшихся в окружающей среде. Примером древнего способа обмена веществ, дошедшего до наших дней, является гликолиз — ферментативное бескислородное расщепление глюкозы. По мере истощения запаса органического материала возникала жесткая конкурентная борьба за него, что ускорило процесс эволюции первичных гетеротрофов. Таким образом, первичными организмами нашей планеты были анаэробы (2, 5–3 млрд лет назад). Исключительным событием стало возникновение фотосинтеза у анаэробных бактерий. Фотосинтез освободил клетки от зависимости доступности органики абиогенного происхождения. Побочным продуктом фотосинтеза являлся кислород, накопление которого в атмосфере привело к коренному изменению хода эволюции. Появление озонового экрана защитило первичные организмы от смертельного УФО и положило конец абиогенному синтезу органики. Первые аэробные бактерии появились 2, 4–2, 8 млрд лет назад благодаря приобретению аппарата окислительного фосфорилирования. Продукты брожения подвергались дальнейшему окислению до СО2 и Н2О. Аэробные (вторичные) гетеротрофы могли более эффективно, чем анаэробные (первичные) гетеротрофы, расщеплять органические вещества, образующиеся в результате фотосинтеза (рис. 61).

Рис. 61. Эволюция пробионтов
С ростом концентрации кислорода в атмосфере усложнялась жизнь первичных анаэробных гетеротрофов. Некоторые из них вымерли, другие нашли бес-кислородную среду (метанобразующие бактерии или серные бактерии, живущие в горячих подземных источниках). Некоторые первичные гетеротрофы пошли по пути, приведшему к образованию эукариотических клеток. Часть из них вступила в симбиоз с аэробными бактериями, способными к окислительному фосфорилированию. Поглотив вторичных гетеротрофов, первичные не расщепили их на молекулы, а сохранили в качестве энергетических станций, называемых сегодня митохондриями. Такие симбионты дали начало царствам животных и грибов. Другая часть первичных гетеротрофов «заключила союз» не только с аэробными гетеротрофами, но и с первичными фотосинтетиками, сохранив последних в качестве хлоропластов. Такие симбионты дали начало царству растений. В пользу симбиотической теории образования эукариот говорят наличие двух мембран у митохондрий и хлоропластов (внутренняя — своя, наружная — образована клеткой-захватчиком) и идеальный генетический код, позволивший эукариотам отстранится от чужой генетической информации. Митохондрии и хлоропласты имеют кольцевую ДНК и бактериальные рибосомы потому, что их предки были бактериями. Сравнительный биохимический анализ показывает, что в основе энергетического обмена всех без исключения организмов лежат одни и те же поразительно сходные между собой цепи реакций, не связанных с потреблением свободного кислорода, — реакции, которые происходят в клетках современных анаэробов.
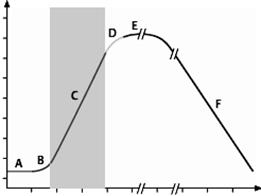
РОСТ И РАЗМНОЖЕНИЕ МИКРООРГАНИЗМОВ РОСТ Рост микроорганизмов — генетически контролируемое увеличение объема и массы микробной клетки, связанное с синтезом новых веществ. Рост в применении к популяции — увеличение биомассы популяции. Стадии (фазы) роста бактериальной культуры на питательной среде (рис. 62). Каждая фаза роста культуры в питательной среде характеризуется определенным размером клеток, скоростью размножения и потребления субстрата, синтезом метаболитов. A — фаза задержки роста (начальная стационарная) , или лаг-фаза (от англ. lag — отставание), в среднем длится 1–2 ч. Начало лаг-фазы связано с адаптацией клеток к среде обитания. Важную роль играет «предыстория» выращивания посевной культуры. Если использован инокулят из культуры с резко отличающимися условиями выращивания, то клеткам требуется время на синтез новых рибосом, РНК и адаптивных ферментов. В этом периоде увеличивается размер клеток, в 8–12 раз повышается содержание РНК. Деления клеток при этом почти не происходит. Полноценная среда, физиологически активная посевная культура, которая подготовлена к синхронному делению, способствуют короткой лаг-фазе (или ее отсутствию) и переходу ко II фазе. Синхронизации можно достичь с помощью пониженной температуры, ограничения питательных веществ, фильтрации, обеспечивающей пропускание клеток определенного размера. Синхронизация длится 2–4 генерации, а далее наступает асинхронный рост.
Время Рис. 62. Фазы роста бактериальной культуры в питательной среде: A — лаг-фаза; B — период положительного ускорения; C — лог-фаза; D — фаза отрицательного ускорения; E — стационарная фаза; F — фаза отмирания B — короткий период положительного ускорения между фазами A и B, когда начинается деление бактерии. C — фаза логарифмического (экспоненциального) роста начинается, когда скорость роста клеток всей популяции достигает постоянной величины, средняя продолжительность ее 5–6 ч. Скорость деления клеток максимальная, но клетки имеют наименьший размер. Популяция бактериальной культуры состоит из делящихся клеток и достаточно стандартна по своим свойствам (содержание белка, нуклеиновых кислот, наиболее выраженные видовые признаки), поэтому эта фаза удобна для определения многих параметров популяции (плотность бактерий, скорости роста и потребления субстрата, содержание биополимеров клетки). В этот период отмечено снижение резистентности к агрессивным веществам. Несмотря на постоянную скорость роста популяции бактерий в логарифмической фазе, отдельные клетки все же находятся в разных стадиях деления. Иногда важно синхронизировать рост всех клеток популяции, то есть получить синхронную культуру. Простыми методами синхронизации являются изменение температурных условий или культивирование в условиях недостатка питательных веществ. Вначале культуру помещают в неоптимальные условия, затем сменяют их оптимальными. При этом у всех клеток популяции синхронизируется цикл деления, но синхронное деление клеток происходит обычно не более 3–4 циклов. D — фаза замедления скорости роста (отрицательного ускорения) длится около 2 ч. Количество питательных веществ существенно уменьшается (отмечается воздействие на бактерии лимитирующих факторов), в культуральной жидкости накапливаются метаболиты, в т. ч. токсичные для бактерий (отмечается ингибирующее воздействие) и скорость деления клеток снижается. E — стационарная фаза, или фаза максимальной концентрации (М-кон-центрация). Клетки перестают делиться. Однако количество живых клеток постоянно, т. к. количество жизнеспособных бактерий соизмеримо с количеством отмирающих. В этот период клетки переходят на эндогенные субстраты (окисляют запасные вещества, белки, углеводы, липиды). Длительность стационарной фазы различается у разных микроорганизмов. Например, у E. coli она наступает через 18–24 ч, у Azotobacter — через 72 ч с момента внесения инокулята в питательную среду. F — фаза отмирания , характеризуется массовой гибелью бактерий. В бактериальной популяции отмечается образование инволюционных форм, аутолиз под действием собственных ферментов. У бактерий меняются морфологические и биохимические свойства. Гибель может наступить через несколько дней, недель, месяцев. В эту фазу различают периоды ускоренной гибели (количество живых клеток начинает снижаться с увеличивающейся скоростью), логарифмической гибели (количество живых клеток убывает с максимальной скоростью), уменьшения скорости гибели (количество живых клеток убывает с уменьшающейся скоростью) и стационарную фазу минимума (количество живых клеток минимально). Данная динамика характерна для периодических (статических) культур с постепенным истощением запаса питательных веществ и накоплением метаболитов. Таким образом, неограниченный рост в закрытой от доступа дополнительных питательных веществ периодической культуре невозможен. Если в питательной среде создают условия для поддержания микробной популяции в экспоненциальной фазе — это хемостатные (непрерывные) культуры. РАЗМНОЖЕНИЕ Размножение микроорганизмов — увеличение их концентрации в единице объема среды, направленное на сохранение вида. Для микроорганизмов характерно: разнообразие способов размножения; переключение с одного способа размножения на другой; возможность одновременного использования нескольких способов; высокая скорость размножения. Способы размножения микроорганизмов: I. Половой способ размножения наблюдается только у эукариот. |
Последнее изменение этой страницы: 2019-04-09; Просмотров: 399; Нарушение авторского права страницы