|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ФІЗІОЛОГІЯ ЗБУДЛИВИХ ТКАНИНСтр 1 из 9Следующая ⇒
ФІЗІОЛОГІЯ ЗБУДЛИВИХ ТКАНИН План
1. Основні властивості збудливих тканин ср 1.1. Подразники. Класифікація подразників ср 1.2. Показники виміру збуджуваності 1.3. Теорія Ходжкіна -Хакслі 2. Мембранний потенціал спокою 3. Мембранний потенціал дії 3.1. Зміни збудливості клітини під час збудження 3.2. Іонні канали 4. Біоструми ср 4.1. Лабільність 4.2. Ритми подразнення 4.3. Парабіоз 5. Фізіологія нервів 5.1. Будова нервової системи. М’якотні та безм’ְ якотні нервові волокна ср 5.2. Властивості нервових волокон 6. Фізіологія м'язів 6.1. Гістологічна будова скелетних м’язів ср 6.2. Основні властивості скелетних м’язів 6.3. Теорія м’язового скорочення 6.4. Робота м’язів 7. Синапси
Теорія Ходжкіна - Хакслі Розберемо розвиток збудження на прикладі нервового волокна. Теорій, що пояснюють розвиток процесу збудження багато, але ми зупинимося на одній з останніх, експериментально найбільше підтвердженої. Це теорія Ходжкина - Хакслі, або теорія калієво - натрієвого насоса. Відповідно до цієї теорії в мембрані клітин, товщиною біля 75А ( IА - 10-8 ) є велика кількість пор і каналів діаметром 7 - 8 А. Пори несуть визначений електричний заряд, що полегшує або утруднює дифузію іонів через мембрану. Мембрана більш проникна для катіонів, ніж для аніонів. Різні катіони також проходять через мембрану з неоднаковою швидкістю. За допомогою мікроелектродної техніки встановлено, що електричний потенціал на мембрані позитивний, а усередині - негативний. Різниця потенціалів пояснюється іонним складом усередині і поза клітиною. Різниця в іонному складі пояснюється насамперед неоднаковим розподілом іонів натрію і калію - так у цитоплазмі нервових м'язових клітин концентрація калію майже в 40 разів вище, чим зовні, а натрію - у 12 разів вище зовні, чим усередині. Для простоти можна вважати, що іони натрію знаходяться зовні клітини, а іони калію - усередині. При цьому спеціальні системи активно переносять іони калію через мембрану усередину клітини і викидають іони натрію, що просочилися в результаті дифузії в клітину з неї.
Звичайно, цей опис будови і функції клітинної мембрани дуже спрощений, але він дає нам можливість роздивитися, як виникає і поширюється нервовий імпульс, тобто хвиля збудження. Початковою реакцією клітини на подразнення є зміна проникності клітинної мембрани в місці подразнення. Іони натрію миттєво поступають усередину клітини, вносячи позитивний заряд. У цей же час із клітини виходить незначна кількість іонів калію. За один імпульс клітина втрачає мільйонну частину всієї кількості калію. У результаті іонних переміщень наступає деполяризація - клітинний потенціал падає до нуля. Потім відбувається перезарядження мембрани - її реполяризація. Через якийсь час потенціал мембрани відновляється і клітина знову може реагувати на подразнення. Просування іонів натрію усередину клітини є причиною появи місцевого кругового електричного струму, що поширюється по поверхні мембрани. Усередині клітини круговий струм йде від збудженого місця до незбудженого, а зовні - від місця спочинку до збудженої ділянки. Проникаючи в сусідню незбуджену ділянку, місцевий струм відчиняє пори для натрію, створюючи тут новий імпульс. Зароджуючись у кожній такій точці нервового або м'язового волокна, імпульс поширюється на значну відстань. Нервовий імпульс - складний електрохімічний процес. Для переміщення іонів калію і натрію проти градієнту концентрації необхідно використання енергії. Ця енергія утворюється в результаті розщеплення молекул АТФ. Жива клітина при здійсненні своїх функцій безперервно генерує і підтримує електричні потенціали, тому універсальним способом з'ясування механізмів діяльності збудливих структур є електрофізіологічні дослідження. Основними іонами, які беруть доля в генерації електричних потенціалів клітин, є К+, Nа+ , Са2+ і Сl–. За умовами виникнення в живих структурах розрізняють мембранні потенціали спокою (МПС) та потенціали дії (ПД).
Мембранний потенціал спокою Мембрана клітини електрично поляризована, і різницю потенціалів між зовнішньою і внутрішньою пластинками (поверхнями) клітинної мембрани називають мембранним потенціалом спокою (МПС). Для того щоб зрозуміти природу виникнення цього потенціалу, треба розглянути деякі властивості самої мембрани, а також склад поза- і внутрішньоклітинної рідин. Клітинна мембрана має значний опір, який значною мірою залежить від стану її йонних каналів і ємність, яка створюється переважно її ліпідним матриксом, і є досить сталою. Так, у аксона кальмара питомий опір клітинної мембрани (Rм) становить у стані спокою 1000 Ом•см2, а ємність (См) — приблизно 1 мкФ/см2. Існування МПС зумовлене передусім нерівномірною концентрацією іонів у поза- та внутрішньоклітинному просторі. Другим важливим чинником, який має значення для створення МПС, є різна проникливість клітинної мембрани для різних іонів. Клітинна мембрана легко проникна для жиророзчинних речовин, молекули яких проходять крізь її ліпідний матрикс. Водорозчинні молекули великого діаметра, у тому числі й аніони органічних кислот, практично не проникають крізь мембрану У стані фізіологічного спокою клітинна мембрана є проникною лише для К+, оскільки в ній є спеціальні калієві канали, що пропускають лише гідратований калій. Коли К+ виходить з клітини, він затримується на зовнішній поверхні клітинної мембрани, створюючи її позитивний заряд. Великі аніони не можуть пройти з клітини крізь мембрану і накопичуються біля її внутрішньої поверхні, створюючи негативний потенціал. Так виникає трансмембранна різниця потенціалів, або МПС. Цей потенціал протидіє подальшому виходу К+ за електрохімічним градієнтом. Переміщення К+ із клітини назовні зумовлюється його концентраційним градієнтом, який здійснює осмотичну роботу (Аосм). Унаслідок МПС К+ частково повертається назад у клітину. При цьому здійснюється електрична робота(Аел) . . Мал. 1. Будова клітинної мембрани (о), її еквівалентна електрична схема (е) і схема активного перенесення йонів за рахунок енергії АТФ (б): / — іонний насос; 2 — ліпідний матрикс (білкові молекули, то утворюють іонні канали і насос, заштриховані); горизонтальними стрілками показано ворітні механізми, вертикальними — напрямок руху іонів; ЕNа, ЕК, Енас — потенціал натрію, калію, насоса; Rм, См — опір та ємність клітинної мембрани
Якщо вихід К+ з клітини переважає над надходженням, то поступово зростає Аелі дещо зменшується аосм. Внаслідок цього на клітинній мембрані виникає потенціал Е, за якого Аел=аосм для К+ і який називають калієвим рівноважним потенціалом (ЕК). Різницю між поточним значенням МПС і Ек називають електрохімічним градієнтом для КК. Він є основною причиною пасивного руху К+ крізь мембрану в природних умовах. Якщо відношення внутрішньої і зовнішньої концентрації іонів калію становить, наприклад, 39, а температура середовища + 20˚ С то Ек становитиме 0, 092 В. За такого значення Ек різниця концентрацій К+ між внутрішнім і зовнішнім середовищами клітини залишається практично незмінною, незважаючи на високу проникливість клітинної мембрани до К+, оскільки підтримується постійною роботою натрій-калієвого насоса. Слід зауважити, що на підтримання МПС витрачається дуже мало К+, основна їх частина повинна забезпечувати електронейтральність внутрішнього середовища клітини. Адже внутрішньоклітинні аніони – це переважно великі білкові молекули, які не можуть пройти крізь клітинну мембрану, і тому їх концентрація залишається сталою. Ці великі аніони мають бути зрівноважені відповідною кількістю катіонів. У зв'язку з тим, що вміст Nа+ всередині клітини дуже незначний, цю функцію виконують К+. Тому внутрішньоклітинна концентрація К+ має бути такою ж високою, як і концентрація великих аніонів, і такою ж стабільною. МПС створює електричне поле, яке діє на макромолекули мембрани й надає їхнім зарядженим групам певну просторову орієнтацію. Електричне поле МПС також забезпечує функцію йонних каналів клітинної мембрани. У стані спокою клітинна мембрана є добре проникною не лише для К+, вона певною мірою проникна також для інших іонів, зокрема Nа+, С1–, Са2+. Потоки цих іонів порушують рівноважний стан. Тої факт, що в умовах спокою Nа+ постійні надходить у клітину, а К+ виходить з неї, має дуже важливі наслідки, оскільки система не може бути фактично врівноважена, за допомогою звичайної дифузії і простого відновлення заряду на клітинній мембрані. Якщо немає інших процесів, то клітина дуже швидко накопичуватиме Nа+, втрачатиме К+, що зменшить їх трансмембранний градієнт. Отже, цей процес може призвести до різкого зниження МПС і загибелі клітини. Проте такого не відбувається, оскільки цьому протидіє натрій-калієвий насос. Оскільки Nа+ сам не може вийти з клітини, а К+ — надійти в клітину проти своїх електрохімічних градієнтів, вони повинні активно переміщуватися крізь мембрану і на забезпечення цього транспорту має витрачатися енергія. Такий механізм дістав назву натрій-калієвого насоса. Він працює, споживаючи енергію АТФ, і його основним компонентом є фермент мембранна натрій-калій-АТФ-аза. У нормі АТФ потрапляє до насоса з мітохондрій аксоплазми. Для роботи насоса обов'язково мають бути зовні К+, а всередині — Nа+. Енергія розщеплення однієї молекулі АТФ забезпечує виведення з клітині трьох іонів натрію і введення всередині клітини двох іонів калію. Це означає, що натрій-калієвий насос є електрогенним: він створює на клітинній мембрані додаткову різницю потенціалів Енас, яка додається до рівня МПС. Цей електрогенний додаток до нормального рівня МПС неоднаковий для різних клітин. Зазвичай у нервових клітин хребетних він незначний, а в гігантських нейронах молюсків і гладком'язових клітинах може становити до 25 % значенні МПС. У формуванні МПС натрій-каліевий насос виконує дві функції: 1) підтримує трансмембранний градієнт концентрацій Nа+ і К+; 2) генерує певну різницю потенціалів, яка додається до потенціалу що створюється дифузією К+ за концентраційним градієнтом. Важливим показником здатності іонів проходити крізь клітинну мембрану є її проникність . Значення проникності для іонів К+, Nа+ і Сl– (РК РNа, РСl) можна використовувати для обчислення МПС мембрани, проникної для всіх цих іонів, за рівнянням Гольдмана — Ходжкіна— Катца (рівняння постійного поля).
Потенціал дії Потенціалом дії (ПД) називають швидке коливання МПС, що виникає під час збудження клітини. В основі ПД лежать зміни електричної (іонної) провідності клітинної мембрани. Амплітуда і тривалість ПД окремої клітини мало залежать від сили подразнення, важливо лише, щоб ця сила досягла певного критичного значення — порогової сили подразнення . ПД, що виникає у місці подразнення, поширюється вздовж нервового чи м'язового волокна, не змінюючи своєї амплітуди. Наявність порогу подразнення і незалежність амплітуди ПД від сили подразнення підпорядковуються закону " все або нічого". Для реєстрації ПД використовують поза- і внутрішньоклітинні електроди. При позаклітинному відведенні електроди підводять до зовнішньої поверхні клітини (волокна). Реєстрація ПД за допомогою позаклітинних електродів показала, що поверхня збудженої ділянки на дуже короткий час (мілісекунди) стає зарядженою негативно відносно сусідньої незбудженої ділянки. Рис. Мембранна провідність під час розвитку ПД у гігантському аксоні кальмара
Під час слідової деполяризації збудливість клітини підвищується (фаза екзальтації), а під час слідової гіперполяризації — знижується (фаза субнормальності), оскільки значення МПС у цей час зростає, при цьому збільшується і критичний (пороговий) рівень деполяризації мембрани.
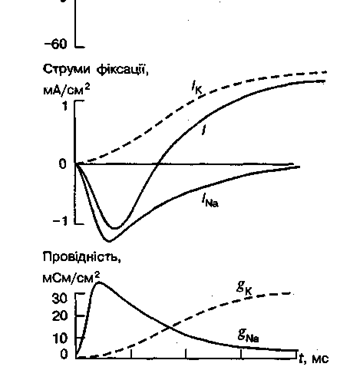
Іонні канали У живій клітині існують дві системи руху іонів крізь мембрану. Одна з них здійснюється за градієнтом концентрації іонів і не потребує витрати енергії — це пасивний іонний транспорт. Він відповідає за підтримання МПС і виникнення ПД, здійснюється через іонні канали. Друга система руху іонів крізь мембрану відбувається проти концентраційного градієнта і полягає у " викачуванні" Na+ з цитоплазми і " закачуванні" К+ всередину клітини за допомогою і онних насосів. Цей механізм можливий тільки з витратами енергії і називається активним іонним транспортом . Функції іонних каналів. У клітинній мембрані існують пори, або канали. Терміном " канал" позначають той шлях, яким іони проходять крізь мембрану за електрохімічним градієнтом. У клітинній мембрані існує кілька різних іонних каналів: селективних та неселективних . Перші пропускають лише якийсь певний іон: Na+, К+ або Са2+, а другі — кілька іонів. Крім того, селективні канали поділяють на потенціалозалежні, потенціалонезалежні (хемочутливі) і канали витоку. Потенціалозалежні канали. Провідність цих каналів контролюється рівнем МПС, тобто ці канали відкриваються (активуються) і закриваються (інактивуються) у відповідь на зміни МПС. Потенціалозалежні канали функціонують в електрозбудливих мембранах і беруть участь у генерації ПД. Електрична збудливість пов’язана з існуванням у таких каналах особливого молекулярного пристрою — воріт, відкривання і закривання яких визначається електричним полем мембрани. Деполяризація мембрани, мВ
Мал. Іонні струми і зміна провідності при зміні МП у гігантському аксоні кальмара: зверху показано деполяризаційне зрушення потенціалу від -60 до 0 мВ. Верхні криві показують підсумковий струм фіксації I та його компоненти Iк і INa; нижні криві показують часовий перебіг змін мембранної провідності gNа і gк, розрахованих за рівнем струмів фіксації
Ворота можуть перебувати тільки у двох положеннях: повністю відчиненими чи повністю зачиненими, тому провідність поодинокого іонного каналу є сталою. Через відкриті канали іони рухаються за концентраційним і електрохімічним градієнтами. Ці потоки іонів зумовлюють зміну МПС, що, в свою чергу, змінює середню кількість відкритих каналів і відповідно рівень іонних струмів. Для дослідження функції іонних каналів застосовують різні методи. Один із них це метод фіксації напруги на мембрані за допомогою якого на мембрані штучно підтримується будь-який потрібний потенціал. Функцію каналів відбивають іонні струми, які в цей час проходять крізь мембрану. Потенціалозалежними є натрієві, калієві і кальцієві канали. Натрієві і кальцієві канали відповідають за ранній (початковий) струм вхідного (в клітину) напрямку, а калієві — за пізній (відставлений) струм вихідного (з клітини) напрямку. Рівень gNа і gк показує кількість одночасно відкритих натрієвих і калієвих каналів. Як можна побачити, gNа за частки мілісекунди досягає максимуму, а потім повільно знижується до початкового рівня. Це пояснюється наявністю у натрієвих каналах двох типів воріт — швидких активаційних і повільних інактиваційних . Отже, початкове збільшення gNа пов'язане з відкриванням перших воріт (процес активації), а наступне повільне зниження gNа, що відбувається під час деполяризації мембрани, зумовлене: закриванням других воріт (процес інактктивації). Натрієвим каналам властива досить висока, але не абсолютна вибірковість Nа+, оскільки більшою або меншою вони є проникними також для іонів літію, талію і навіть деяких органічних катіонів (гідразин, амоній). Отвір натрієвого каналу має прямокутну форму розмірами 0, 3х0, 3 нм. Вузька частина каналу, що є селективним фільтром, дуже коротка і має одиничний негативний заряд. Цей заряд притягує катіони, відштовхує аніони, забезпечуючи вибіркову проникність для Nа+ і подібних до нього катіонів.
а б в Мал. Потенціалозалежний натрієвий канал: Внутрішня частина каналу (1), утвореного макромолекулою білка (2), — це селективний фільтр; т — активаційні. h — інактиваційні: - ворота каналу; а — стан спокою (активаційні ворота зачинені, інактнваційні — відчинені); б — початок деполяризації мембрани (швидке відчинення активаційних воріт); в — інактивація каналу (інактиваційні ворота зачинені); поверхня клітинної мембрани: А — зовнішня; Б — цитоплазматична
Активаційні та інактиваційні ворота розміщені біля внутрішнього кінця натрієвого каналу, причому інактиваційні ворота спрямовані в бік цитоплазми. У стані спокою активаційні ворота закриті, а інактиваційні відкриті. Під час деполяризації мембрани спочатку відкриті обидва типи воріт, тобто канал пропускає Nа+. Потім інактиваційні ворота повільно закриваються — канал інактивується. Лише після закінчення деполяризації поступово відкриваються іиактиваційні ворота, а активаційні закриваються і канал повертається до початкового стану. Деякі речовини специфічно змінюють процеси активації та інактивації натрієвих каналів. Так, батрахотоксин спричинює стійку деполяризацію клітинної мембрани, підвищуючи gNa. Він повністю усуває процес натрієвої інактивації так, що натрієві канали стають постійно відкритими. Тетродотаксин дуже швидко і різко пригнічує ранній іонний струм крізь мембрану, але тільки коли він діє із зовні, тобто на чутливі до тетродотоксину рецепторні структури натрієвих каналів, розміщені на зовнішній поверхні мембрани. Калієві канали за будовою подібні до натрієвих, але відрізняються своєю вибірковістю, спрямованістю потоку іонів, кінетикою процесів інактивації і активації. В усіх збудливих клітинах калієві канали відповідають за пізній іонний струм. Діаметр калієвого каналу у найвужчій його частині становить 0, 3 нм. Отже, калієві канали проникні для катіонів розміром 0, 26-0, З нм (К+, RЬ+). Активація цих каналів порівняно з натрієвими відбувається досить повільно. Крім того, протягом перших 10 мс деполяризації немає ніякої калієвої інактивації, вона виникає тільки під час дуже тривалої (кілька секунд) деполяризації мембрани. Викладене вище про співвідношення між процесами активації та інактивації калієвих каналів властиве лише нервовим волокнам. У мембрані багатьох нервових і м'язових клітин існують калієві канали, які інактивуються порівняно швидко. Виявлено також калієві канали з швидкою активацією. Нарешті, є калієві канали, які активуються не мембранним потенціалом, а внутрішньоклітинним Са2+. Щільність розміщення калієвих каналів на мембрані дещо менша, ніж натрієвих. Специфічним блокатором калієвих каналів є тетраетиламоній (ТЕА), і они гідрогену, амінопіридини. ТЕА діє з будь-якого боку клітинної мембрани. Кальцієві канали. Ретельне дослідження функції іонних каналів різних нервових клітин дало змогу модифікувати іонну модель генерації ПД, запропоновану Ходжкіним і Хакслі, до якої входили лише натрієві й калієві канали. Виявилося, що Са2+ також беруть активну участь у генерації ПД. З'ясувалося також, що в деяких клітинах вхідний струм створюється переважно Са2+ (гладком'язові клітини). Крім порівняно швидкого вхідного кальцієвого струму, що бере участь у генерації ПД, було виявлено ще один дуже повільний кальцієвий струм. Цей струм відповідає за повільну деполяризацію (протягом кількох секунд), яка зумовлює генерацію серій імпульсів деяких пейсмекерних клітин, наприклад серцевих провідних міоцитів (клітин Пуркіньє). Селективність (вибірковість) кальцієвих каналів зумовлена наявністю в ділянці зовнішнього входу хімічних груп, які мають підвищену спорідненість до двовалентних катіонів: іони кальцію зв'язуються з такими групами і тільки після цього проходять до порожнини каналу. До деяких двовалентних катіонів, наприклад Мn2+, спорідненість цих хімічних груп така велика, що, зв'язуючись з ними, ці катіони блокують рух Са2+ через канал. Характерною особливістю кальцієвих каналів є залежність їх від клітинного метаболізму, зокрема від циклічних нуклеотидів (цАМФ, цГМФ), які регулюють процеси фосфоритування і дефосфоритування білків кальцієвих каналів. Іони кальцію ефективно впливають на синтез білків і забезпечують транспорт утворених молекул до аксонів і дендритів. Відкриття цього явища безпосередньо свідчило про те, що процеси на мембрані прямо пов'язані з процесами всередині клітини. Дослідження останніх років показали, що Са2+ є важливим регулятором активності збудливих систем. Переважна кількість Са2+, що потрапляє в клітини під час збудження, зв'язується буферними системами цитозолю, мітохондріями та іншими внутрішньоклітинними депо. Проте та невелика частина Са2*, що залишається вільною, бере активну участь у таких процесах, як регуляція функції К+ і Сl— каналів, Са2+, Nа+ обміну тощо. Виявлено ще одну особливість кальцієвих каналів: вони здатні дуже специфічно реагувати на деякі фізіологічні речовини, що утворюються іншими клітинами і виділяються у внутрішнє середовище організму (катехоламіни, пептиди тощо). Під впливом цих речовин канали активуються чи, навпаки, інактивуються Внаслідок цього змінюється активність нервової клітини. Потенціалонезалежні (хемочутливі) канали. Активність цих каналів контролюється тільки хеморецепторами. Активація хеморецепторів медіаторами супроводжується здебільшого відкриванням каналів. Хемочутливі потенціалонезалежні канали функціонують у субсінаптичних мембранах і беруть участь у: генерації синоптичних потенціалів. Вони реагують не тільки на нейромедіатори, а й на гормони та деякі інші фізіологічно активні речовини. Канали витоку також потенціалонезалежні, але вони не контролюються хеморецепторами. Ці канали відіграють істотну роль у сумарній провідності мембран у стані спокою, саме їхньою активністю пояснюють менші значення МПС порівняно з розрахованим за рівнянням. Функцію цих каналів ще не з’ясовано. Кожний потенціалозалежний іонний канал навіть за сталих значень МПС то відкривається, то закривається. Це відбувається незакономірно, тобто процес має ймовірний (стохастичний) характер. Таке явище викликає переміщення іонів і створює електричний шум. Аналіз цих шумів дав змогу розрахувати щільність розміщення іонних каналів у мембрані і провідність одного каналу. Так, у гігантському аксоні кальмара щільність натрієвих каналів становить 300 на 1 мкм2, середня провідність –4 пСм (пікосименсів); у вузлах нервового волокна(перетяжках Ранв'є) щільність натрієвих каналів значно вища — 2000 на 1 мкм; а провідність удвічі більша — 8 пСм. _
Біоструми Виходячи з усього вищевикладеного, ви можете легко зрозуміти відкіля беруться біоструми в живій тканині і чому вони відсутні в мертвих клітинах. Історія вчення про біоструми нараховує вже декілька сторіч. У 1791 році італійський фізіолог Луіджі Гальвані довів, що в живій тканині є біоструми. Наявність біострумів він довів дослідами, які ввійшли в історію за назвою 1 і 2 дослідів Гальвані. Він установив наявність, так називаних, токів покою або токів ушкодження. Крім того, під час збудження на поверхні мембрани реєструються струми дії. Між ними є розходження. Якщо струми ушкодження однофазні і реєструються довгостроково, то токи дії двофазні і короткочасні. Лабільність У зв'язку зі зміною потенціалу дії, а отже і з тривалістю проходження фаз збуджуваності, знаходиться і функціональна рухливість або лабільність тканини. Вчення про лабільність створив російський учений Н. Е. Введенський. Для визначення частоти виникаючих потенціалів дії, він у 1884 році застосував слухавку, сполучену з нервом нервово - м'язового препарату. При подразненні нерва з різноманітною частотою в слухавці виникали звуки - тони різноманітної висоти. Чим частіше завдавалося подразнення, тим вище був тон. По граничній висоті тону можна було судити про максимальну кількість відтворених у секунду імпульсів збудження. Таке ж положення спостерігалося і при подразненні м'язів. Н. Е. Введенський створив вчення про функціональну рухливість, або лабільність, що розкриває закономірності ритмічного збудження, що поширюється Основним у вченні про лабільність є положення про те, що в кожній тканині одиночний імпульс збудження продовжується визначений час. Ритми подразнення При ритмічному подразненні визначеної частоти тканина не відразу відповідає відповідною кількістю імпульсів збудження. Це залежить від того, що лабільність до початку подразнення відрізняється від лабільності під час подразнення - вона змінюється в зв'язку з діяльністю. Дія подразника і виникаючі імпульси збудження змінюють швидкість процесів обміну речовин у тканині. Остання набуває спроможності відтворювати більш частий, раніше недоступний їй ритм. Дослідами Н. Е. Введенського вперше було встановлено, що найвище скорочення м'яза спостерігається при нанесенні на нерв нервово - м'язового препарату подразнень у більш рідкому ритмі. Такий найбільш вигідний у робочому відношенні ритм подразнення називається оптимальним. При оптимальному ритмі подразнень кожний новий імпульс збудження виникає під час фази екзальтації, створеної попереднім імпульсом, у результаті чого і відбувається максимальне скорочення м'яза. Оптимальний ритм порушень менше максимального в декілька разів. Наприклад, у жаби максимальний ритм для рухового нерва складає 500, а оптимальний - 100 - 150 імпульсів у секунду, для литкового м'яза відповідно - 120 - 200 і 30 - 50 імпульсів у секунду. При дуже частих подразненнях, що перевищують оптимальний ритм, скорочення м'яза зменшується і навіть зовсім припиняється - спостерігається песимум ритму подразнення або песимум частоти. Песимум спостерігається в тому випадку, коли частота подразнень перевищує міру лабільності. Під впливом першого подразнення тканина збуджується, імпульс збудження ще не закінчився, а вже завдається повторне подразнення. Повторне подразнення діє під час фази абсолютної рефрактерності. Хоча те подразнення не в змозі викликати збудження, воно усе ж не залишається без наслідків - поглиблює фазу абсолютної рефрактерності і затрудняє дію наступних подразнень. Тому кожне нове подразнення, що посилається в дуже частому ритмі, знижує лабільність тканини і викликає не збудження, а якісно новий стан - гальмування. Явища оптимуму і песимуму, відкриті Н. Е. Введенським при вивченні реакцій нервово - м'язового препарату, виявилися загальнобіологичними властивостями, що спостерігаються в живій тканині у відповідь на будь-які фізичні або хімічні подразники. Встановлено, що помірні подразнення забезпечують оптимальну (збудливу) дію, а подразнення великої сили, частоти і тривалості забезпечують песимальну (гальмуючу) дію на реакції різноманітних органів як при безпосередньому, так і при рефлекторному впливі на них.
Парабіоз Явища оптимуму і песимуму відзначаються не тільки в рухових реакціях, але й у реакціях секреторних органів і сердечно - судинної системи. Ці явища були виявлені й в умовно-рефлекторних реакціях різноманітної складності. Вивчаючи вплив різноманітних тривалих хімічних і фізичних подразнень на нерв нервово - м'язового препарату, Н. Е. Введенський установив закономірні стадії зміни функціонального стана нерва в подразнюючийся ділянці. Виходячи з цих досліджень він сформулював теорію про парабіоз ( паро - біля, біос - життя ). Ця теорія пояснює, як збудження переходить у гальмування, а гальмування може закінчитися припиненням життєдіяльності тканини, тобто її смертю. Фізіологічна сутність парабіозу була в основному розкрита Н. Е. Введенським; остаточне роз'яснення вона одержала в роботах його учнів і послідовників, що застосували новітні методи фізіологічних досліджень. Н. Е. Введенський діяв на середню частину нерва різноманітними хімічними речовинами, викликаючи зміни (альтерирування) даної ділянки. До дії речовини, що альтерирує, відповідь м'яза залежала від сили і частоти подразнення: чим більше сила або частота, тим вище скорочення м'яза. При впливі на нерв речовиною, що альтерирує (викликає зміни), через якийсь час відповідь м'яза на подразники різноманітної сили і частоти змінилися. Фізіологія нервів
5.1. Будова нервової системи. М’якотні та безмְ ’якотні нервові волокна Нервова система утворена нейронами, що являють собою нервову клітину з відростками, різними за розміром, формою і числу. Відросток, що проводить збудження від клітини називається аксоном, а відросток, що проводить збудження до клітини зветься дендритом. Відросток нервової клітини складається з осьового циліндра, покритого оболонками, разом із якими він утворюєь нервове волокно. Розрізняють м’якотні і безм’якотні нервові волокна. У м’якотних волокнах є мієлінова оболонка. У окремих ділянках волокна, так званих перехопленнях Ранвье, мієлінова оболонка переривається. М’якотні волокно входять до складу більшості спинномозкових нервів. У безм’якотних волокнах навколо осьових циліндрів немає мієлінової оболонки. З безм’якотних волокон утворені постганглионі волокна вегетативної нервової системи. М’якотні і безм'якотні нервові волокна йдуть пучками, декілька пучків складають нервовий стовбур, або нерв. Одні з нервових волокон проводять збудження від периферії до нервових центрів - це аферентні, або доцентрові, волокна; інші проводять збудження від центрів на периферію - це еферентні, або відцентрові, волокна. Більшість нервів є змішаними, тому що в їхньому складі йдуть і аферентні і еферентні волокна.
5.2. Властивості нервових волокон:
1. Збудливість. Фізіологія м'язів. Аеробна фаза (ЦТК) Молочна кислота → СО2 і Н2О Приблизно 1/5 частина молочної кислоти утворюється з глікогену, а інші 4/5 знову перетворюються у глікоген).
М’язи деякий час можуть скорочуватися без кисню, але потім зупиняються, тому що використовується АТФ, креатин фосфат і гексозо фосфат, накопичується молочна кислота, для окислення якої необхідний кисень. Кількість кисню, необхідна для окислення молочної кислоти, називається кисневим запитом. При скороченні м’язів утворюється багато енергії. Приблизно 30% перетворюється в механічну, а остаток виділяється у вигляді тепла у дві фази: 1. на початку скорочення – 40% 2. при розслабленні – 60%
Робота м’язів Сила м’яза виявляється в його максимальному напруженні. М’язову силу можна вимірювати за допомогою динамометрів — приладів, що реєструють напруження м’яза, який перебуває в умовах ізометричного скорочення. Сила м’яза залежіть від товщини, тобто фізіологічного поперечника, який визначають як суму площ розтину всіх м’язових волокон, перерізаних перпендикулярно до їх довжини. Тому сила м’язів різної анатомічної будови буде різна. Найбільшу силу мають перисті м’язи, у яких фізіологічний поперечник перевищує анатомічній. Також сила м’язів залежіть від початкової довжини м’яза. Розтягнутий м’яз здатен підняти значно більший вантаж порівняно з нерозтягнутим. На силу м’язів впливає функціональний стан організму. М’язова сила змінюється при тренуванні та з віком. Вона зменшується при старінні, голодуванні та стомленні. Для порівняння сили м’язів введено поняття абсолютної сили. Абсолютна сила – величина найбільшого вантажу, який м’яз може підняти в перерахунку на 1 см2 фізіологічного поперечника. Абсолютна сила вимірюється в кілограмах на 1 см2. Абсолютна сила триголового м'яза плеча людини – 16, 8 кг/см2., м’язів жаби – 2 – 3, а м'язів комах – 6, 9 кг/см2. При підйомі вантажу м’яз виконує механічну роботу, що вимірюється добутком маси вантажу (P)на висоту підйому (h).
W = Ph. Остільки величина роботи залежить від маси вантажу та висоти скорочення м’яза, важливим фактором, що впливає на висоту скорочення, є довжина м’язових волокон. М’язи з довгими, паралельно розташованими волокнами піднімають вантаж на більшу висоту, ніж перисті. Попереднє помірне розтягування м’яза збільшує висоту його скорочення. При надмірному розтягненні вона помітно зменшується. Дослідами доведено, що ізольований м’яз виконує найбільшу роботу при середньому навантаженні. При інтенсивній роботі м’яза працездатність зменшується. Такий стан назувають втомою. Синапси Складне утворення, що забезпечує перехід збудження з нервового волокна на м'язову, нервову або залозисту клітину називається синапсом. Всі синапси поділяють на центральні і периферичні. Периферичний синапс - це область контакту між аксоном і працюючим органом, Центральний синапс - область контакту аксона одного нейрона з дендритом або нервовою клітиною іншого нейрона. Синапс складається з трьох основних елементів - пресипатичної мембрани, постсинаптичної мембрани і синоптичної щілини. Проведення збудження через синапс є складним процесом. Після дослідів О. Леви, що установили роль хімічних речовин - медіаторів - у передачі збудження з нервів на серце, стали вважати, що збудження через синапси проводяться медіаторами. А. Ф. Самойлов ( 1925 ) установив, що передача збудження з нерва на м'яз прискорюється при підвищенні температури. Він також вважав, що збудження через синапси передається за допомогою медіаторів. Наявність медіаторів збудження в міжнейронних синапсах, як-от у синапсах симпатичної нервової системи, встановив А. В. Кибяков ( 1923 ). Пізніше було відкрито, що медіатором, за допомогою якого передається збудження в нервово - м'язових скелетних і міжнейронних синапсах, є ацетилхолін. Закінчення постганглионарних симпатичних нервових волокон звільняють в основному норадреналін і незначну кількість адреналіну. Ацетилхолін не тільки бере участь у передачі збудження, він також впливає на процеси обміну речовин і обумовлює підвищення лабільності, необхідне для більш швидкого проведення наступних імпульсів збудження. В даний час установлено, що в механізмі передачі збудження через синапси одночасно беруть участь і електричний струм і хімічні речовини. Імпульс збудження звільняє ацетилхолін із пухирців нервового закінчення. Ацетилхолін дифундує через пресинаптичну мембрану в симпатичну щілину і викликає деполяризацію постсинаптичної мембрани й у ній з'являється постсинаптичний потенціал дії. Під впливом його виникають кругові токи, що збуджують сусідні ділянки м'язового волокна. Ацетилхолін, що виділився, дуже швидко руйнується ферментом холінестеразою на холін і оцтову кислоту. Руйнація ацетилхоліну відбувається протягом рефракторного періоду, що викликається попереднім імпульсом збудження. Тому кожна порція ацетилхоліну, що виділився, діє дуже короткотривало (біля 0, 001 секунди), встигаючи викликати тільки 1 імпульс. Синапси проводять збудження в одну сторону - від нервового волокна до м'язового. Одностороннє проведення збудження обумовлене тим, що медіатор виділяється тільки в нервовому закінченні. Збудження через синапси проводиться значно повільніше, чим у нервах. Це уповільнення називається синоптичною затримкою. Вона обумовлена особливостями передачі збудження через синапс і залежності від часу, необхідного для дифузії медіатора через пресинаптичну мембрану і синоптичну щілину до постсинаптичної мембрани і для виникнення під впливом медіатора постсинаптичного потенціалу дії. Синапси мають малу збуджуваність і низьку лабільність, то гальмування насамперед наступає в синусах.
Література
1. Георгиевский В.И. Физиология сельскохозяйственных животных. – М.: Агропромиздат, 1990. – 511 с. 2. Науменко В.В. та ін. Фізіологія сільськогосподарських тварин: Підручник. – Київ: Вид-во " Сільгоспосвіта", 1994. – 512 с. 3. Воронцов Д.С., Ємченко А.І. Фізіологія тварин і людини. – К.: Радянська школа, 1952. – 655 с. 4. Кіндя В.І., Куровський Ю.А., Мусієнко В.Ф. Словник-довідник з анатомії та фізіології с.-г. тварин. – К.: " Урожай", 1993. – 294 с. 5. Рихтер и др. Основные физиологические показатели у животных и технология содержания. – М.: Колос, 1982. – 192 с. 6. Шмидт-Ниельсен К. Физиология животных. Приспособление и среда. Книги 1, 2. – М.: Мир, 1982. – 800 с. 7. Шмидт-Ниельсен К. Размеры животных: почему они так важны? – М.: Мир, 1987. – 259 с. 8. Физиология сельскохозяйственных животных.-М.: Агропромиздат, 1991. - 432 с. 9. Общий курс физиологии человека и животных. В 2 кн. Кн. 2. Физиология висцеральных систем.- М.: Высш. Шк., 1991. - 528 с. ФІЗІОЛОГІЯ ЗБУДЛИВИХ ТКАНИН План
1. Основні властивості збудливих тканин ср 1.1. Подразники. Класифікація подразників ср 1.2. Показники виміру збуджуваності 1.3. Теорія Ходжкіна -Хакслі 2. Мембранний потенціал спокою 3. Мембранний потенціал дії 3.1. Зміни збудливості клітини під час збудження 3.2. Іонні канали 4. Біоструми ср 4.1. Лабільність 4.2. Ритми подразнення 4.3. Парабіоз 5. Фізіологія нервів 5.1. Будова нервової системи. М’якотні та безм’ְ якотні нервові волокна ср 5.2. Властивості нервових волокон 6. Фізіологія м'язів 6.1. Гістологічна будова скелетних м’язів ср 6.2. Основні властивості скелетних м’язів 6.3. Теорія м’язового скорочення 6.4. Робота м’язів 7. Синапси
|
Последнее изменение этой страницы: 2019-06-08; Просмотров: 350; Нарушение авторского права страницы