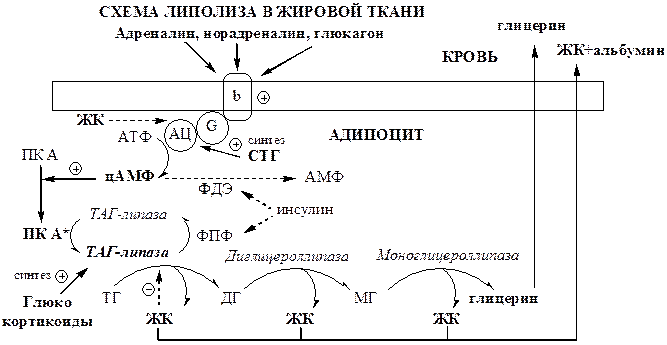|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Тема: Переваривание и всасывание липидов. Транспорт липидов в организме.Стр 1 из 11Следующая ⇒
Кафедра биохимии
КУРС ЛЕКЦИЙ ПО ОБЩЕЙ БИОХИМИИ для студентов 2 курса лечебно-профилактического факультета
Модуль 4. Биохимия липидов
Автор: к.б.н., доцент кафедры биохимии Гаврилов И.В.
Екатеринбург, 2013г Тема: Переваривание и всасывание липидов. Транспорт липидов в организме. Обмен липопротеидов. Дислипопротеидемии.
Липиды - это разнообразная по строению группа органических веществ, которые объединены общим свойством - растворимостью в неполярных растворителях. КЛАССИФИКАЦИЯ ЛИПИДОВ Липиды по способности к гидролизу делят на омыляемые (двух и более компонентные) и неомыляемые (однокомпонентные). Омыляемые липиды в щелочной среде гидролизуются с образованием мыл, они содержат в своем в составе жирные кислоты и спирты глицерин (глицеролипиды) или сфингозин (сфинголипиды). По количеству компонентов омыляемые липиды делятся на простые (состоят из 2 классов соединений) и сложные (состоят из 3 и более классов). К простым липидам относятся: 1) воска (сложный эфир высшего одноатомного спирта и жирной кислоты); 2) триацилглицериды, диацилглицериды, моноацилглицериды (сложный эфир глицерина и жирных кислот). У человека весом в 70 кг ТГ около 10 кг. 3) церамиды (сложный эфир сфингозина и жирной кислоты С18-26) – лежат в основе сфинголипидов; К сложным липидам относятся: 1) фосфолипиды (содержат фосфорную кислоту): а) фосфоглицеролипиды (сложный эфир глицерина и 2 жирных кислот, содержит фосфорную кислоту и аминоспирт) - фосфатидилсерин, фосфатидилэтаноламин, фосфатидилхолин, фосфатидилинозитол, фосфатидилглицерол; б) кардиолипины (2 фосфатидные кислоты, соединенные через глицерин); в) плазмалогены (сложный эфир глицерина и жирной кислоты, содержит ненасыщенный одноатомный высший спирт, фосфорную кислоту и аминоспирт) – фосфатидальэтаноламины, фосфатидальсерины, фосфатидальхолины; г) сфингомиелины (сложный эфир сфингозина и жирной кислоты С18-26, содержит фосфорную кислоту и аминоспирт - холин); 2) гликолипиды (производные сфингозина, содержащие углеводы): а) цереброзиды (сложный эфир сфингозина и жирной кислоты С18-26, содержит гексозу: глюкозу или галактозу); б) сульфатиды (сложный эфир сфингозина и жирной кислоты С18-26, содержит гексозу (глюкозу или галактозу) к которой присоединена в 3 положение серная кислота). Много в белом веществе; в) ганглиозиды (сложный эфир сфингозина и жирной кислоты С18-26, содержит олигосахарид из гексоз и сиаловых кислот). Находятся в ганглиозных клетках; К неомыляемым липидам относят: 1. стероиды; 2. жирные кислоты (структурный компонент омыляемых липидов), 3. витамины А, Д, Е, К; 4. терпены (углеводороды, спирты, альдегиды и кетоны с несколькими звеньями изопрена). БИОЛОГИЧЕСКИЕ ФУНКЦИИ ЛИПИДОВ В организме липиды выполняют разнообразные функции: 1) Структурная. Сложные липиды и холестерин амфифильны, они образуют все клеточные мембраны; фосфолипиды выстилают поверхность альвеол, образуют оболочку липопротеинов. Сфингомиелины, плазмалогены, гликолипиды образуют миелиновые оболочки и другие мембраны нервных тканей. 2) Энергетическая. В организме до 33% всей энергии АТФ образуется за счет окисления липидов; 3) Антиоксидантная. Витамины А, Д, Е, К препятсвуют СРО; 4) Запасающая. ТГ являются формой хранения жирных кислот; 5) Защитная. ТГ, в составе жировой ткани, обеспечивают теплоизоляционную и механическую защиту тканей. Воска образуют защитную смазку на коже человека; 6) Регуляторная. Фосфотидилинозитолы являются внутриклеточными посредниками в действии гормонов (инозитолтрифосфатная система). Из полиненасыщенных жирных кислот образуются эйкозаноиды(лейкотриены, тромбоксаны, простагландины, простациклины), вещества, регулирующие иммуногенез, гемостаз, неспецифическую резистентность организма, воспалительные, аллергические, пролиферативные реакции. Из холестерина образуются стероидные гормоны: половые, кортикоиды, кальцитриол; 7) Пищеварительная. Из холестерина синтезируются желчные кислоты. Желчные кислоты, фосфолипиды, холестерин обеспечивают эмульгирование и всасывание липидов; 8) Информационная. Ганглиозиды обеспечивают межклеточные контакты. Источником липидов в организме являются синтетические процессы и пища. Некоторые липиды в организме не синтезируются (полиненасыщенные жирные кислоты - витамин F, витамины А, Д, Е, К), они являются незаменимыми и поступают в организм только с пищей. ПРИНЦИПЫ НОРМИРОВАНИЯ ЛИПИДОВ В ПИТАНИИ В сутки человеку требуется потреблять 80-100г липидов, из них 25-30г растительного масла, 30-50г сливочного масла и 20-30г жира животного происхождения. Потребность в пищевых липидах зависит от возраста. Новорожденным до 3 месяцев требуется 6, 5 г/кг липидов, детям до 6 месяцев - 6 г/кг, детям после 6 месяцев – 5, 5 г/кг, взрослым – 1, 4 г/кг, пожилым – 0, 5 г/кг. Причины: 1). основным источником энергии для детей грудного возраста являются липиды, а для взрослых людей - глюкоза. 2). Энергозатраты с возрастом снижаются. Потребность в липидах увеличивается на холоде, при физических нагрузках, в период выздоровления и при беременности. С пищей в норме поступает около 85-90г ТГ, 1г ФЛ, 0, 3—0, 5 г ХС (в основном в виде эфиров). Растительные масла содержат много полиеновых незаменимых (линолевая до 60%, линоленовая) жирных кислот, фосфолипидов (удаляются при рафинировании). Сливочное масло содержит много витаминов А, Д, Е. Все природные липиды хорошо перевариваются, масла усваиваются лучше жиров. При смешанном питании сливочное масло усваивается на 93-98%, свиной жир - на 96-98%, говяжий жир – на 80-94%, подсолнечное масло – на 86-90%. Длительная тепловая обработка (> 30 мин) разрушает полезные липиды, при этом образуются токсические продукты окисления жирных кислот и канцерогенные вещества. При недостаточном поступлении липидов с пищей снижается иммунитет, снижается продукция стероидных гормонов, нарушается половая функция. При дефиците линолевой кислоты развивается тромбоз сосудов и увеличивается риск раковых заболеваний. При избытке липидов в пище развивается атеросклероз и увеличивается риск рака молочной железы и толстой кишки. ПЕРЕВАРИВАНИЕ ЛИПИДОВ Переваривание – это гидролиз пищевых веществ до их ассимилируемых форм. Лишь 40-50% пищевых липидов расщепляется полностью, от 3% до 10% пищевых липидов всасываются в неизмененном виде. Так как липиды не растворимы в воде, их переваривание и всасывание имеет свои особенности и протекает в несколько стадий: 1) Липиды твердой пищи при механическом воздействии и под влиянием ПАВ желчи смешиваются с пищеварительными соками с образованием эмульсии (масло в воде). Образование эмульсии необходимо для увеличения площади действия ферментов, т.к. они работают только в водной фазе. Липиды жидкой пищи (молоко, бульон и т.д.) поступают в организм сразу в виде эмульсии; 2) Под действием липаз пищеварительных соков происходит гидролиз липидов эмульсии с образованием водорастворимых веществ и более простых липидов; 3) Выделенные из эмульсии водорастворимые вещества всасываются и поступают в кровь. Выделенные из эмульсии более простые липиды, соединяясь с компонентами желчи, образуют мицеллы; 4) Мицеллы обеспечивают всасывание липидов в клетки эндотелия кишечника.
Ротовая полость В ротовой полости происходит механическое измельчение твердой пищи и смачивание ее слюной (рН=6, 8). У грудных детей здесь начинается гидролиз ТГ с короткими и средними жирными кислотами, которые поступают с жидкой пищей в виде эмульсии. Гидролиз осуществляет лингвальная триглицеридлипаза («липаза языка», ТГЛ), которую секретируют железы Эбнера, находящиеся на дорсальной поверхности языка. Желудок Так как «липаза языка» действует в диапазоне 2-7, 5 рН, она может функционировать в желудке в течение 1-2 часов, расщепляя до 30% триглицеридов с короткими жирными кислотами. У грудных детей и детей младшего возраста она активно гидролизует ТГ молока, которые содержат в основном жирные кислоты с короткой и средней длиной цепей (4—12 С). У взрослых людей вклад «липазы языка» в переваривание ТГ незначителен. В главных клетках желудка вырабатывается желудочная липаза, которая активна при нейтральном значении рН, характерном для желудочного сока детей грудного и младшего возраста, и не активна у взрослых (рН желудочного сока ~1, 5). Эта липаза гидролизует ТГ, отщепляя, в основном, жирные кислоты у третьего атома углерода глицерола. Образующиеся в желудке ЖК и МГ далее участвуют в эмульгировании липидов в двенадцатиперстной кишке. Тонкая кишка Основной процесс переваривания липидов происходит в тонкой кишке. 1. Эмульгированиелипидов (смешивание липидов с водой) происходит в тонкой кишке под действием желчи. Желчь синтезируется в печени, концентрируется в желчном пузыре и после приёма жирной пищи выделяется в просвет двенадцатиперстной кишки (500-1500 мл/сут). Жёлчь это вязкая жёлто-зелёная жидкость, имеет рН=7, 3-8.0, содержит Н2О – 87-97%, органические вещества (желчные кислоты – 310 ммоль/л (10, 3-91, 4 г/л), жирные кислоты – 1, 4-3, 2 г/л, пигменты желчные – 3, 2 ммоль/л (5, 3-9, 8 г/л), холестерин – 25 ммоль/л (0, 6-2, 6) г/л, фосфолипиды – 8 ммоль/л) и минеральные компоненты (натрий 130-145 ммоль/л, хлор 75-100 ммоль/л, НСО3- 10-28 ммоль/л, калий 5-9 ммоль/л). Нарушение соотношение компонентов желчи приводит к образованию камней. Жёлчные кислоты (производные холановой кислоты) синтезируются в печени из холестерина (холиевая, и хенодезоксихолиевая кислоты) и образуются в кишечнике (дезоксихолиевая, литохолиевая, и д.р. около 20) из холиевой и хенодезоксихолиевой кислот под действием микроорганизмов.
В желчи желчные кислоты присутствуют в основном в виде конъюгатов с глицином (66-80%) и таурином (20-34%), образуя парные желчные кислоты: таурохолевую, гликохолевую и д.р.
Соли жёлчных кислот, мыла, фосфолипиды, белки и щелочная среда желчи действуют как детергенты (ПАВ), они снижают поверхностное натяжение липидных капель, в результате крупные капли распадаются на множество мелких, т.е. происходит эмульгирование. Эмульгированию также способствует перистальтика кишечника и выделяющийся, при взаимодействии химуса и бикарбонатов СО2: Н+ + НСО3- → Н2СО3 → Н2О + ↑ СО2. 2. Гидролизтриглицеридов осуществляет панкреатическая липаза. Ее оптимум рН=8, она гидролизует ТГ преимущественно в положениях 1 и 3, с образованием 2 свободных жирных кислот и 2-моноацилглицерола (2-МГ). 2-МГ является хорошим эмульгатором. 28% 2-МГ под действием изомеразы превращается в 1-МГ. Большая часть 1-МГ гидролизуется панкреатической липазой до глицерина и жирной кислоты. В поджелудочной железе панкреатическая липаза синтезируется вместе с белком колипазой. Колипаза образуется в неактивном виде и в кишечнике активируется трипсином путем частичного протеолиза. Колипаза своим гидрофобным доменом связывается с поверхностью липидной капли, а гидрофильным способствует максимальному приближению активного центра панкреатической липазы к ТГ, что ускоряет их гидролиз.
3. Гидролизлецитина происходит с участием фосфолипаз (ФЛ): А1, А2, С, D и лизофосфолипазы (лизоФЛ).
В результате действия этих четырех ферментов фосфолипиды расщепляются до свободных жирных кислот, глицерола, фосфорной кислоты и аминоспирта или его аналога, например, аминокислоты серина, однако часть фосфолипидов расщепляется при участии фосфолипазы А2 только до лизофосфолипидов и в таком виде может поступать в стенку кишечника. ФЛ А2 активируется частичным протеолизом с участием трипсина и гидролизует лецитин до лизолецитина. Лизолецитин является хорошим эмульгатором. ЛизоФЛ гидролизует часть лизолецитина до глицерофосфохолина. Остальные фосфолипиды не гидролизуются. 4. Гидролизэфиров холестерина до холестерина и жирных кислот осуществляет холестеролэстераза, фермент поджелудочной железы и кишечного сока. 5. Мицеллообразование Водонерастворимые продукты гидролиза (жирные кислоты с длинной цепью, 2-МГ, холестерол, лизолецитины, фосфолипиды) вместе с компонентами желчи (солями жёлчных кислот, ХС, ФЛ) образуют в просвете кишечника структуры, называемые смешанными мицеллами. Смешанные мицеллы построены таким образом, что гидрофобные части молекул обращены внутрь мицеллы (жирные кислоты, 2-МГ, 1-МГ), а гидрофильные (желчные кислоты, фосфолипиды, ХС) — наружу, поэтому мицеллы хорошо растворяются в водной фазе содержимого тонкой кишки. Стабильность мицелл обеспечивается в основном солями жёлчных кислот, а также моноглицеридами и лизофосфолипидами.
Пища стимулирует секрецию из клеток слизистой тонкой кишки в кровь холецистокинина (панкреозимин, пептидный гормон). Он вызывает выделение в просвет двенадцатиперстной кишки желчи из желчного пузыря и панкреатического сока из поджелудочной железы. Кислый химус стимулирует секрецию из клеток слизистой тонкой кишки в кровь секретина (пептидный гормон). Секретин стимулирует секрецию бикарбоната (НСО3-) в сок поджелудочной железы. ОБМЕН ЛИПИДОВ В ЭНТЕРОЦИТАХ Липиды поступают в энтероциты как из просвета кишечника, так и из тканей. Большая часть липидов, поступивших в энтероцит, подвергается ресинтезу. 1. 1-МГ гидролизуется кишечной липазой до глицерина и жирной кислоты. 2. Короткоцепочечные жирные кислоты, ФЛ (кроме лецитина) и часть глицерина без изменений направляются из энтероцита в кровь. 3.Длинноцепочечные эндогенные и экзогенные жирные кислоты под действием ацил-КоА-синтетазы (тиокиназы) активируются, образуя Ацил~КоА: RCOOH + HS-КоА + АТФ → Ацил~КоА + АМФ + ФФн Обмен липопротеинов Липопротеины (ЛП) – это надмолекулярные комплексы сферической формы, состоящие из липидов, белков и углеводов. ЛП имеют гидрофильную оболочку и гидрофобное ядро. В гидрофильную оболочку входят белки и амфифильные липиды - ФЛ, ХС. В гидрофобное ядро входят гидрофобные липиды - ТГ, эфиры ХС и т.д. ЛП хорошо растворимы в воде.
В организме синтезируются несколько видов ЛП, они отличаются химическим составом, образуются в разных местах и осуществляют транспорт липидов в различных направлениях. ЛП разделяют с помощью: 1) электрофореза, по заряду и размеру, на α -ЛП, β -ЛП, пре-β -ЛП и ХМ; 2) центрифугирования, по плотности, на ЛПВП, ЛПНП, ЛППП, ЛПОНП и ХМ. Соотношение и количество ЛП в крови зависит от времени суток и от питания. В постабсорбтивный период и при голодании в крови присутствуют только ЛПНП и ЛПВП. Основные виды липопротеинов
Апобелки Белки, входящие в состав ЛП, называются апопротеины (апобелки, апо). К наиболее распространенным апопротеинам относят: апо А-I, А-II, В-48, В-100, С-I, С-II, С-III, D, Е. Апобелки могут быть периферическими (гидрофильные: А-II, С-II, Е) и интегральными (имеют гидрофобный участок: В-48, В-100). Периферические апо переходят между ЛП, а интегральные – нет. Апопротеины выполняют несколько функций:
Ферменты транспорта липидов Липопротеинлипаза(ЛПЛ) (КФ 3.1.1.34, ген LPL, около 40 дефектных аллелей) связана с гепарансульфатом, находящимся на поверхности эндотелиальных клеток капилляров кровеносных сосудов. Она гидролизует ТГ в составе ЛП до глицерина и 3 жирных кислот. При потере ТГ, ХМ превращаются в остаточные ХМ, а ЛПОНП повышают свою плотность до ЛППП и ЛПНП. Апо С-II ЛП активирует ЛПЛ, а фосфолипиды ЛП участвуют в связывании ЛПЛ с поверхностью ЛП. Синтез ЛПЛ индуцируется инсулином. Апо С-III ингибирует ЛПЛ. ЛПЛ синтезируется в клетках многих тканей: жировой, мышечной, в легких, селезёнке, клетках лактирующей молочной железы. Ее нет в печени. Изоферменты ЛПЛ разных тканей отличаются по значением Кm. В жировой ткани ЛПЛ имеет Кm в 10 раз больше, чем в миокарде, поэтому в жировая ткань поглощает жирные кислоты только при избытке ТГ в крови, а миокард – постоянно, даже при низкой концентрации ТГ в крови. Жирные кислоты в адипоцитах используются для синтеза ТГ, в миокарде как источник энергии. Печёночная липаза находиться на поверхности гепатоцитов, она не действует на зрелые ХМ, а гидролизует ТГ в ЛППП. Лецитин: холестерол-ацил-трансфераза (ЛХАТ) находиться в ЛПВП, она переносит ацил с лецитина на ХС с образование ЭХС и лизолецитина. Ее активируют апо А-I, А-II и С-I. лецитин + ХС → лизолецитин + ЭХС ЭХС погружается в ядро ЛПВП или переноситься с участием апо D на другие ЛП. НОРМАЛЬНЫЕ ЗНАЧЕНИЯ В соответствии c рекомендациями ASSMANN в настоящее время следующие значения холестерина ЛПВП могут быть предложены как основополагающие для оценки риска развития атеросклероза:
ОБМЕН ХИЛОМИКРОНОВ Липиды, ресинтезированные в энтероцитах, транспортируется тканям в составе ХМ.
· Образование ХМ начинается с синтеза апо В-48 на рибосомах. Апо В-48 и В-100 имеют общий ген. Если с гена копируется на мРНК только 48% информации, то с нее синтезируется апо В-48, если 100% - то с нее синтезируется апо В-100. · С рибосом апо В-48 поступает в просвет ЭПР, где он гликозилируется. Затем в аппарате Гольджи апо В-48 окружается липидами и происходит формирование «незрелых», насцентных ХМ. · Экзоцитозом насцентные ХМ выделяются в межклеточное пространство, поступают в лимфатические капилляры и по лимфатической системе, через главный грудной лимфатический проток попадают в кровь. · В лимфе и крови с ЛПВП на насцентные ХМ переносятся апо Е и С-II, ХМ превращаются в «зрелые». ХМ имеют довольно большой размер, поэтому они придают плазме крови опалесцирующий, похожий на молоко, вид. Под действием ЛПЛ ТГ ХМ гидролизуются на жирные кислоты и глицерол. Основная масса жирных кислот проникает в ткань, а глицерол транспортируется с кровью в печень. · Когда в ХМ количество ТГ снижается на 90%, они уменьшаются в размерах, а апо С-II переносится обратно на ЛПВП, «зрелые» ХМ превращаются в «остаточные» ремнантные ХМ. Ремнантные ХМ содержат в себе фосфолипиды, холестерол, жирорастворимые витамины и апо В-48 и Е. · Через ЛПНП-рецептор (захват апо Е, В100, В48) ремнантные ХМ захватываются гепатоцитами. Путём эндоцитоза остаточные ХМ попадают внутрь клеток и перевариваются в лизосомах. ХМ исчезают из крови в течение нескольких часов.
ОБМЕН ЛПВП ЛПВП выполняют 2 основные функции: они поставляют апо другим ЛП в крови и участвуют в так называемом «обратном транспорте ХС». ЛПВП синтезируются в печени и в небольшом количестве в тонком кишечнике в виде насцентных ЛПВП. Они имеют дисковидную форму, небольшой размер и содержат высокий процент белков и фосфолипидов. В печени в ЛПВП включаются апопротеины А, Е, С-II, ЛХАТ. В крови апо С-II и апо Е переносятся с ЛПВП на ХМ и ЛПОНП. насцентные ЛПВП практически не содержат ХС и ТГ и в крови обогащаются ХС, получая его из других ЛП и мембран клеток. Для переноса ХС в ЛПВП существует сложный механизм. На поверхности ЛПВП находится фермент ЛХАТ — лецитин: холестерол-ацилтрансфераза. Этот фермент превращает ХС в ЭХС. Реакция активируется апо A-I, входящим в состав ЛПВП. ЭХС перемещается внутрь ЛПВП. Таким образом, ЛПВП обогащаются ЭХС. ЛПВП увеличиваются в размерах, из дисковидных небольших частиц превращаются в частицы сферической формы, которые называют ЛПВП3, или «зрелые ЛПВП». ЛПВП3 частично обменивают ЭХС на ТГ, содержащиеся в ЛПОНП, ЛППП и ХМ. В этом переносе участвует «белок, переносящий эфиры холестерина» - апо D. Таким образом, часть ЭХС переносится на ЛПОНП, ЛППП, а ЛПВП3 за счёт накопления ТГ увеличиваются в размерах и превращаются в ЛПВП2. Часть ЛПВП захватывается клетками печени, взаимодействуя со специфическими для ЛПВП рецепторами к апо А-1. На поверхности клеток печени ФЛ и ТГ ЛППП, ЛПВП2 гидролизуются печёночной липазой, что дестабилизирует структуру поверхности ЛП и способствует диффузии ХС в гепатоциты. ЛПВП2 в результате этого опять превращаются в ЛПВП3 и возвращаются в кровоток. НАРУШЕНИЯ ЛПВП Болезнь Тэнжи Болеют аборигены острова Тэнжи. Наследственный дефект апо А, не синтезируются ЛПВП. Нарушается транспорт излишков ХС из тканей в печень. В крови низкий уровень ХС, ФЛ, много ТГ. Макрофаги фагоцитируют в тканях излишки ХС с образованием ксантом. Накопление ХС в печени, селезенке и других лимфоидных органах вызывает гепатоспленомегалию и лимфаденопатию. Может развиваться катаракта, полинейропатия и ренит. Миндалины из-за отложений ХС окрашены в оранжево-желтый цвет. Список литературы Берсенёв Алексей Вячеславович. Кандидатская диссертация: Трансплантация клеток эмбриональной печени и стволовых клеток костного мозга для коррекции дислипидемии и ранних стадий атерогенеза. М.: 2003.
ЛЕКЦИЯ № 13 Развитие жировой ткани
ХИМИЧЕСКИЙ СОСТАВ БЕЛОЙ ЖИРОВОЙ ТКАНИ Жировая ткань содержит 65-85% ТГ, 22% воды, 5, 8% белка, 15 ммоль/кг калия. Из жирных кислот 42—51% приходится на олеиновую, 22—31% - на пальмитиновую, 5—14% - на пальмитоолеиновую, 3—5 % - на миристиновую, 1—5 % - на линолевую кислоты. Состав жировой ткани зависит от области тела, глубины слоя; он может также несколько отличаться у отдельных индивидуумов. Особенно подвергается изменениям содержание воды и белка. Чем глубже под поверхностью кожи жир расположен, тем больше он содержит насыщенных кислот. У новорожденных насыщенные жиры во всех слоях содержатся в одинаковом количестве. ОСОБЕННОСТИ МЕТАБОЛИЗМА БЕЛОЙ ЖИРОВОЙ ТКАНИ Энергетический обмен низкий, преимущественно анаэробный, ткань потребляет мало кислорода. Энергия АТФ в основном тратится на транспорт жирных кислот через клеточные мембраны (с участием карнитина). Белковый обмен низкий, белки синтезируются адипоцитами преимущественно для собственных нужд. На экспорт в жировой ткани синтезируются лептин, белки острой фазы воспаления (α 1-кислый гликопротеин, гаптоглобин), компоненты системы комплимента (адипсин, комплемент С3, фактор В), интерлейкины. Углеводный обмен. Невысокий, преобладает катаболизм. Углеводный обмен в жировой ткани тесно связан с липидным. Липидный обмен Жировая ткань стоит на 2 месте по обмену липидов после печени. Здесь происходят реакции липолиза и липогенеза. Липогенез. В жировой ткани синтез липидов идет в абсорбтивный период по глицерофосфатному пути. Процесс стимулируется инсулином. Этапы липогенеза: 1. Под действием инсулина на рибосомах стимулируется синтез ЛПЛ. 2. ЛПЛ выходит из адипоцита и фиксируется на поверхности стенки капилляра с помощью гепарансульфата. 3. ЛПЛ гидролизует ТГ в составе липопротеинов
4. Образовавшийся глицерин уноситься кровью в печень. 5. Жирные кислоты из крови транспортируются в адипоцит. 6. Кроме поступающих из вне экзогенных жирных кислот, в адипоците синтезируются жирные кислоты из глюкозы. Процесс стимулируется инсулином. 7. Жирные кислоты в адипоците под действием Ацил-КоА синтетазы превращаются в Ацил-КоА.
7. Глюкоза поступает в адипоцит с участием ГЛЮТ-4 (активатор инсулин). 8. В адипоците глюкоза вступает в гликолиз с образованием ФДА (активатор инсулин). 9. В цитоплазме ФДА восстанавливается глицерол-ф ДГ до глицерофосфата:
Так как в жировой ткани нет глицерокиназы, глицерофосфат образуется только из глюкозы (не может из глицерина). 10. В митохондриях глицерофосфат под действием глицеролфосфат ацилтрансферазы превращается в лизофосфатид:
11. В митохондриях лизофосфатид под действием лизофосфатид ацилтрансферазы превращается в фосфатид:
11. Фосфатид под действием фосфотидат фосфогидролазы превращается в 1, 2-ДГ:
12. 1, 2-ДГ под действием ацилтрансферазы превращается в ТГ:
13. Молекулы ТГ объединяются в крупные жировые капли.
В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.
ОСОБЕННОСТИ МЕТАБОЛИЗМА БУРОЙ ЖИРОВОЙ ТКАНИ Энергетический обмен. Ткань потребляет много кислорода, активно окисляет глюкозу и жирные кислоты. Энергетический обмен высокий. При этом, АТФ образуется только в реакциях субстратного фосфорилирования (2 реакции гликолиза, 1 реакции ЦТК). Причина - разобщение в митохондриях белком термогенином (РБ-1) процессов окисления и фосфорилирования, низкая активность АТФ синтетазы, отсутствие дыхательного контроля со стороны АДФ. В бурой жировой ткани вся образующаяся при окислении энергия рассеивается в виде тепла (термогенез). Термогенез в бурой жировая ткань активируется при переохлаждении СНС, а также при излишке липидов в крови, под действием лептина. Благодаря этому повышается температура тела и снижается концентрация липидов в крови. Отсутствие бурой жировой ткани у взрослых людей является причиной 10% всех случаев ожирения. ЛЕКЦИЯ № 14 Строение жирных кислот Жирными кислотами (ЖК) - называются карбоновые кислоты, которые образуются при гидролизе омыляемых липидов. В основном к жирным кислотам относятся высшие карбоновые кислоты (содержащие 12 и более атомов С). Такие ЖК водонерастворимыми, они транспортируются в крови с помощью альбуминов, а в клетках - с помощью Z-белков. ЖК человека и животных имеют некоторые особенности строения: 1) они монокарбоновые; 2) содержат четное количество атомов С, самая распространенная длина от 16 до 18 атомов С; 3) углеродный скелет неразветвлен; 4) бывают насыщенными и ненасыщенными (мононенасыщенными и полиненасыщенными); 5). двойные связи несопряжены (разделены метиленовыми мостиками) и имеют цис-конформацию.
∆ ЖК – номера атомов С, у которых расположены двойные связи. ω ЖК - число атомов С от последней двойной связи до конца цепи. Биологическое значение ЖК
КАТАБОЛИЗМ ЖИРНЫХ КИСЛОТ В живых организмах катаболизм ЖК протекает как в ферментативных так и в неферментативных реакциях. · Ферментативный катаболизм ЖК происходит в основном в реакциях β -окисления. К побочным путям относиться ферментативное α - и ω -окисление ЖК, а также деградация ЖК в пероксисомах. Хотя эти побочные пути количественно менее важны, их нарушение может приводить к тяжелым заболеваниям. · Неферментативный катаболизм ЖК протекает в реакциях перекисного окисления липидов (ПОЛ). β -окисление ЖК β -окисление — специфический путь катаболизма ЖК с неразветвленной средней и короткой углеводородной цепью. β -окисление протекает в матриксе митохондрий, при котором от С конца ЖК последовательно отделяется по 2 атома С в виде Ацетил-КоА. β -окисление ЖК происходит только в аэробных условиях и является источником большого количества энергии. β -окисление ЖК активно протекает в красных скелетных мышцах, сердечной мышце, почках и печени. ЖК не служат источником энергии для нервных тканей, так как ЖК не проходят через гематоэнцефалический барьер, как и другие гидрофобные вещества. β -окисление ЖК увеличивается в постабсорбтивный период, при голодании и физической работе. При этом концентрация ЖК в крови увеличивается в результате мобилизации ЖК из жировых ткани. Активация ЖК Активация ЖК происходит в результате образования макроэргической связи между ЖК и HSКоА с образованием Ацил-КоА. Реакцию катализирует фермент Ацил-КоА синтетаза: RCOOH + HSKoA + АТФ → RCO~SКоА + АМФ+ PPн Пирофосфат гидролизуется ферментом пирофосфатазой: Н4Р2О7 + Н2О → 2Н3РО4 Ацил-КоА синтетазы находятся как в цитозоле (на внешней мембране митохондрий), так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к ЖК с различной длиной углеводородной цепи. Транспорт ЖК |
Последнее изменение этой страницы: 2017-04-12; Просмотров: 642; Нарушение авторского права страницы




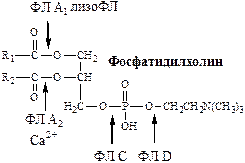
 Регуляция переваривания
Регуляция переваривания 










 2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.