|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Нейронные сети кишечнополостныхСтр 1 из 8Следующая ⇒
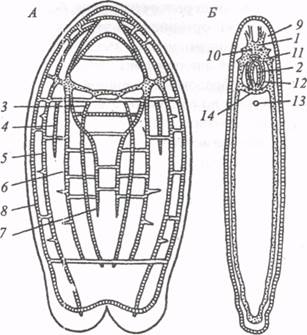
Ортогон плоских червей Как известно, плоские черви - первые трехслойные многоклеточные животные. У них в дополнение к эктодерме и энтодерме появляется третий зародышевый слой - мезодерма, участвующий в формировании различных систем органов (выделения, размножения). Поэтому плоские черви оказываются на более высоком уровне структурной организации. Энтодермальных элементов нервной системы у этих животных нет, и ЦНС выделяется как самостоятельный отдел. Нервный аппарат низших турбеллярий представляет собой диффузное субэпителиальное нервное сплетение, без стволов и продольных сгущений. По своей организации это сплетение находится на уровне нервной сети кишечнополостных. У высших турбеллярий нервный аппарат развит значительно лучше. Эволюционное преобразование нервной системы в пределах типа плоских червей идет по следующим направлениям: 1) централизация нервного аппарата; 2)погружение его элементов в глубь тела; 3)усиление роли головного ганглия; 4)усложнение внутренней структуры, обеспечивающее большую сложность поведения животного. Централизация нервных элементов приводит у высших турбеллярий к преобразованию диффузного нервного сплетения в упорядоченную структуру, называемую ортогоном. Это ассоциативные и двигательные нервные клетки, собранные вместе с отростками в продольные тяжи или стволы, которые соединены множеством поперечных и кольцевых комиссуральных стволов (рис.2). Между перекладинами ортогона остаются только элементы периферической нервной системы, включающие чувствительные клетки и отростки двигательных клеток. Формирование ортогона представляет собой первый шаг к образованию центрального отдела нервной системы. Если в диффузной нервной сети ассоциативные и двигательные клетки нередко удалены друг от друга, то в ортогоне пути между ними укорачиваются. Однако наряду с укорочением внутрицентральных связей наблюдается удлинение связей центра с периферией (с мышцами и чувствительными клетками).
Рис. 2. Нервный аппарат турбеллярий Polychoerus caudatus (А) и Macrostomum finnlandense (Б) (по: В.Н.Беклемишев, 1964): 1 - мозг, 2 - глотка, 3 - внутренние брюшные стволы, 4 - наружные брюшные стволы, 5 - боковые стволы, 6 - наружные спинные стволы, 7 - внутренние спинные стволы, 8 - краевой нерв, 9 - отходящие от мозга чувствительные нервы, 10 - глаза, 11 - нервы глоточного сплетения, 12 - глоточное нервное сплетение, 13 - выделительное отверстие, 14 - передняя комиссура брюшных продольных стволов В пределах типа плоских червей наблюдается тенденция к уменьшению числа стволов и комиссур, т. е. к дальнейшей централизации. Чем выше систематическое положение какой-то группы плоских червей, тем сильнее выражена у особей этой группы концентрация нервных элементов. Например, у турбеллярий Macrostomum finnlandense ортогон состоит только из брюшных продольных стволов и двух поперечных комиссур: передней и каудальной (рис.2). Следует отметить, что нервные клетки в стволах ортогона еще не сгруппированы в ганглии. Возникновение билатеральной симметрии у плоских червей с четко выраженной продольной осью тела и его передним и задним полюсами способствует централизации нервных элементов. На переднем конце тела плоских червей концентрируются рецепторы, среди которых статоцист в виде углубившегося под эпителий замкнутого пузырька с известковыми включениями, «глазки» в форме пигментных пятен или чашечек, обонятельные ямки и множество эпителиальных чувствительных клеток. Огромное число волокон от этих рецепторов приводит к значительной концентрации нервной ткани на переднем конце тела и способствует формированию мозгового, или церебрального, ганглия (эндона), контролирующего другие центры (рис.2). Ассоциативные клетки эндона посылают длинные отростки в продольные стволы ортогона, в связи с чем, прилежащие к мозгу участки продольных стволов резко утолщаются. Таким образом, церебральный ганглий приобретает связи с частями тела, и его роль интегративная возрастает. О регуляторной функции эндона говорят результаты целого ряда физиологических экспериментов на планариях, в первую очередь опытов с удалением мозга. Удаление церебрального ганглия сказывается на «спонтанных» ритмических движениях планарий. Животные одних видов, например Planaria torva, сохраняют спонтанную двигательную активность после экстирпации. Однако скорость локомоции значительно снижена. Удлиняется также скрытый период двигательной реакции, вызванной местным механическим раздражением. Особи других видов, например Thysanozoon broechii, после удаления мозга теряют способность к спонтанным движениям, возбудимость у них снижена, и только при сильном раздражении они начинают плавать. У планарий с удаленным эндоном утрачивается способность узнавать пищу, нарушается правильная ориентация по отношению к источнику света (отрицательный фототаксис). Даже если у обезглавленных планарий сохраняется реакция на свет, она не имеет приспособительного значения. У морских поликлад удаление эндона вызывает прекращение ретроградных движений, связанных с пробеганием волны сокращения вдоль тела. Одностороннее разрушение церебрального ганглия может вызвать у поликлад асимметричное распределение мышечного тонуса и манежные движения, направленные в сторону, противоположную оперированной. Следовательно, церебральный ганглий плоских червей выполняет функции регуляции возбудимости и тонуса сократительного аппарата, а также координации приспособительных реакций, возникающих в ответ на изменения экологических факторов. Плоским червям, как и кишечнополостным, свойственно привыкание к воздействию раздражителей. Например, у планарий Dugesia dorotocephala привыкание к вибрации наступает при интервалах между раздражениями не менее 10-15 с. Животные в ходе опыта научаются тормозить свои реакции на действие периодического вибрационного раздражителя. С использованием классических методов И.П. Павлова у планарий изучали динамику выработки электрооборонительного условного рефлекса на свет. Животных помещали в экспериментальную камеру, дно которой было разделено на темное и освещенное поля. На одном из полей планарии получали удар током, который вызывал продольное сокращение их тела. Н.А. Тушмаловой удалось установить, что приобретенные планариями реакции во многом сходны с суммационными рефлексами. Они характеризуются малой стойкостью даже в ходе одного эксперимента и тем более от опыта к опыту. Вместе с тем динамика угасания выработанных реакций свидетельствует о более длительном сохранении следов от предыдущих раздражений у планарий по сравнению с кишечнополостными. Этот факт свидетельствует о специфичности приобретенных реакций планарий, которые, по мнению Н.А. Тушмаловой, можно классифицировать как примитивные, нестойкие условные рефлексы. Истинные условные рефлексы вырабатываются только у высших беспозвоночных (начиная с полихет), головные ганглии которых обладают более совершенной морфофункциональной организацией. Спинальный этап развития В ходе эволюции трубчатая система позвоночных животных имеет несколько критических этапов развития, на протяжении которых возникают новые уровни интеграции в ростральных, филогенетически более молодых отделах мозга. Исходным считается спинальный тип центральной нервной системы, представленный у наиболее древних хордовых – бесчерепных. В составе нервной трубки современного бесчерепного – ланцетника, нет дифференцированного головного мозга. Спинной мозг имеет форму длинного тяжа, состоящего из 64 однородных сегментов, от которых отходят задние (чувствительные) и передние (двигательные) корешки. Они выходят из спинного мозга отдельными стволами. На поперечном срезе спинной мозг ланцетника имеет трапециевидную форму. В центре трапеции проходит спинномозговой канал. Внутреннее строение нервной трубки отличается примитивностью. Четкого разделения на белое и серое вещество не отмечается, однако нервные клетки имеют тенденцию концентрироваться вокруг центрального канала. Белое вещество спинного мозга ланцетника состоит из безмякотных волокон, в основном малого диаметра. Вместе с тем в ростральных (6-11-й сегмент) и каудальных (39-61-й сегмент) отделах нервной трубки располагаются гигантские клетки Родэ (рис. 11), толстые аксоны которых после дугообразного изгиба следуют на значительное расстояние в пределах спинного мозга, формируя проводящие пути, имеющие координационное значение. К клеткам Родэ в составе задних корешков подходят афферентные аксоны от светочувствительных глазков Гессе, которые локализованы вдоль всей нервной трубки, но преимущественно в передней и хвостовой частях тела ланцетника. Каждый глазок Гессе., состоящий из пигментной и ганглиозной клетки и представляющий собой примитивный фоторецептор, реагирует на световые раздражители, свободно распространяющиеся через прозрачные покровы тела. Возбуждение от фоторецепторов через синаптические переключения достигает клеток Родэ и далее адресуется мотонейронам, аксоны которых иннервируют мышечные пластинки ланцетника. Таким образом, происходит замыкание рефлекторной дуги, лежащей в основе поведенческой реакции – отрицательного фототаксиса. Сенсорный аппарат ланцетника представлен помимо глазков Гессе реснитчатыми клетками эпителия, которые реагируют на химические, термические и механические раздражители, в частности на вибрационные колебания водной среды. Афференты от этих рецепторных образований в составе задних корешков проникают в спинной мозг ланцетника.
Рис. 11. Расположение нервных элементов в нервной, трубке ланцетника (по: А. И. Карамян, 1976; А.Л.Поленов, 1971): А – срединная часть и Б– головной конец нервной трубки; 1– задний корешок, 2 – передний корешок, 3 – глазок Гессе, 4 – клетка Родэ, 5 – спинномозговой канал, 6 – инфундибулярный орган В головной части нервной трубки расположена обонятельная ямка, выстланная клетками чувствительного эпителия. Эти клетки снабжены ресничками и, по мнению ряда авторов, обладают хеморецепторной функцией. Однако вопрос о функциях обонятельной ямки остается мало освещенным. В вентральной стенке переднего конца нервной трубки локализован «инфундибулярный орган», который состоит из крупных клеток, обладающих нейросекреторной функцией. Апикальная часть этих клеток достигает расширенной полости спинномозгового канала, базальный же полюс переходит в отросток, идущий к вентральной поверхности нервной трубки (рис. 11). Цитоплазма клеток инфундибулярного органа заполнена мелкими (100-300 нм) гомориположительными секреторными гранулами. Согласно представлениям А.Л. Поленова (1971), описанная выше структура является гомологом нейросекреторных клеток преоптического или супраоптического ядра гипоталамуса других, более высокоорганизованных позвоночных животных. На основании структурной специфичности головного отдела нервной трубки ланцетника некоторые авторы (Ф.В. Овсянников, В.Я. Данилевский) считали эту область зачатком головного мозга – архенцефалоном, регулирующим всю рефлекторную деятельность бесчерепных:. Однако в результате физиологических исследований, где анализировали восприятие и простые формы обучения у ланцетника, установлено, что нет достаточных оснований говорить о специфике головного отдела нервной трубки. Так, например, по данным Б.Ф. Сергеева, перерезка нервной трубки в ее центральной части ни в коей мере не отражается на условнорефлекторных реакциях ланцетника. Они вырабатываются одинаково успешно как при раздражении рецепторов головного отдела, так и при стимуляции хвостовой части тела. Условнорефлекторные реакции формировались при использовании одномодальных раздражителей, например слабого света, создающего освещенность 30 люкс, как условного раздражителя и сильного света (120 люкс) как безусловного подкрепления, После 28-36 сочетаний условного и безусловного раздражителей слабый свет приобретал сигнальное значение и вызывал двигательную реакцию - отрицательного фототаксиса. Вместе с тем при сочетании разномодальных раздражителей (света и электрического тока) условнорефлекторная реакция не вырабатывалась, что свидетельствовало об отсутствии феномена замыкания между нервными элементами, воспринимающими условный и безусловный раздражители. Эти факты говорят о том, что приобретенные реакции ланцетника по своему механизму отличаются от классических условных рефлексов высших позвоночных. Формирование условных реакций у ланцетника протекает по типу повышения возбудимости афферентных и эфферентных систем спинного мозга при повторном раздражении. В данном случае суммирование следов возбуждения с наличным возбуждением приводит к повышению чувствительности (сенсибилизации) ЦНС, которая начинает реагировать на условные раздражители, бывшие до того подпороговыми. Такие индивидуально приобретенные реакции относятся к категории суммационных рефлексов. У ланцетника суммационные рефлексы характеризуются малой стабильностью и сохраняются не более десяти часов. Из всех видов внутреннего торможения у этих животных вырабатывается только угасательное. Таким образом, бесчерепным доступны наиболее простые формы обучения, которое осуществляется по механизму суммационного рефлекса. Эти функции реализуются теми отделами нервной трубки, которые имеют сенсорный вход от рецепторов (глазков Гессе) и моторный выход к сократительному аппарату. Надсегментарных (мозговых) систем интеграции в ЦНС бесчерепных нет, и в целом можно говорить о равнозначности всех отделов нервной трубки в плане восприятия и простых форм обучения. Отсутствие выраженной структурной и функциональной специализации характерно для наиболее древнего, спинального этапа развития ЦНС позвоночных животных, согласно предложенному А.И. Карамяном принципу этапности в историческом развитии ЦНС. Ихтиопсидный этап развития Морфофизиологические преобразования в спинном мозге рыб сопряжены с появлением более сложных форм двигательного поведения. В частности, переход от стереотипных ундулирующих движений круглоротых к локомоции с помощью плавников связан с дальнейшей дифференциацией кожной и мышечно-суставной (проприоцептивной) чувствительности. У хрящевых пластиножаберных рыб в спинальных ганглиях имеются специализированные нейроны для выполнения этих функций. На базе проприоцептивной чувствительности совершенствуются физиологические механизмы рефлекторного сопротивления мышцы ее растяжению, механизмы динамической фиксации суставов. Наблюдается постепенная концентрация серого вещества в окружности центрального канала. Укорачивается путь моторных аксонов внутри спинного мозга, причем каждый мотонейрон иннервирует не миомер (как у круглоротых), а определенную группу мышц плавника. Белое вещество спинного мозга рыб также претерпевает определенную дифференциацию. Нисходящие пути представлены древним ретикулоспинальным трактом. Среди нейронов, образующих этот тракт, особо выделяются маутнеровские клетки, лежащие в продолговатом мозге Гигантские аксоны маутнеровских клеток посылают множество коллатералей к вставочным и моторным нейронам спинного мозга. Раздражение ретикуло-спинального тракта вызывает ипсилатерально сокращения мышц туловища, жаберной крышки, нижней челюсти; в то же время на противоположной стороне туловища наблюдается торможение моторики. У хрящевых пластиножаберных рыб появляются в виде особых пучков вестибуло-спинальный и церебелло-спинальный тракты, которые связывают спинной мозг с центральными аппаратами регуляции движения (вестибулярным аппаратом и мозжечком). Эти связи двусторонние и дополняются восходящим спино-церебеллярным трактом, проходящим в латеральных столбах от уровня шейной часта: спинного мозга. Спиноцеребеллярный тракт рыб достигает корпуса мозжечка и может быть отождествлен с дорсальным мозжечковым трактом Флексига высших позвоночных. Кроме того, группа восходящих волокон вентролатеральных столбов оканчивается в ретикулярной формации продолговатого мозга и частично в крыше среднего мозга. Сравнительно недавно окончания восходящих спинальных волокон обнаружены у акул в таламусе промежуточного мозга. Долго считали, что филогенетически более молодые восходящие пути дорсальных столбов у рыб, как и у миног, представляют внутреннюю эндоспинальную систему и не имеют проекций в стволовых частях мозга. Однако недавно в дорсальных столбах хрящевых и костистых рыб идентифицированы длинные спинальные волокна в области спинобульбарного соединения. Электрофизиологическими методами доказано участие дорсальных столбов у костистых рыб, как и у круглоротых, в проведении импульсации, вызванной тактильным раздражением. Таким образом, в процессе филогенетического развития происходит постепенное расширение связей спинного мозга с различными отделами головного мозга. Непосредственное продолжение спинного мозга - продолговатый мозг у рыб имеет ряд продольных возвышений, содержащих ядра черепномозговых нервов. Среди них следует отметить ядра вестибулолатеральной системы, с помощью которой осуществляется рефлекторная установка головы и тела в пространстве. Дифференциация рецепторов боковой линии, появление у хрящевых рыб третьего полукружного канала в лабиринте сопряжено со значительным увеличением объема ядер вестибуло-латеральной системы. Дорсальное ядро, в котором оканчиваются толстые волокна переднего нерва боковой линии, превращается в целую долю. Одновременно разрастается медиальное ядро, к которому подходят тонкие волокна переднего и заднего нервов боковой линии. В вентральном ядре, куда проецируются окончания восьмого нерва (вестибулокохлеарного), увеличивается число составляющих это ядро клеток, они становятся крупнее. Слуховые функции у рыб выполняют нижняя часть лабиринта внутреннего уха (саккулюс) и в качестве усиливающего воспринимающего прибора плавательный пузырь. Отметим, что пластино-жаберные рыбы перестают реагировать на низкочастотные звуки (500-600 Гц) после перерезки восьмого нерва. Соматическая афферентная зона в продолговатом мозге рыб представлена помимо системы вестибулокохлеарного нерва системой тройничного нерва (пятая пара). Восходящие проекции ядер тройничного нерва направляются к среднему (тектуму, тегментуму), промежуточному и конечному мозгу. Эфференты ядер тройничного нерва адресованы моторным ядрам стволовой части мозга и ретикулярной формации. У акул эфференты ядер тройничного нерва идут в спинной мозг, часть входов направляется в мозжечок. Чувствительные окончания тройничного нерва рыб имеются в слизистой оболочке ротовой полости. Их возбуждение при наполнении полости рта водой активирует инспираторные нейроны дыхательного центра, осуществляющего автоматическую смену фаз дыхательного цикла. В рефлекторной регуляции дыхательных движений принимают участие также афферентные волокна лицевого (седьмая пара) и блуждающего (десятая пара) нервов. Ядра этих нервов формируют висцеросенсорную зону продолговатого мозга. Сюда поступает информация от хеморецепторов, сосредоточенных на поверхности головы, в жабрах, в глотке. Данная система обусловливает восприятие природы химических и пищевых компонентов окружающей среды. Нейроны моторной части ядра блуждающего нерва координируют деятельность пищеварительного тракта и других внутренних органов. В сетчатой (ретикулярной) формации продолговатого мозга рыб выделяют медианную, медиальную и латеральную зоны. В медианной зоне локализованы переднее, среднее и заднее ядра шва, которые описаны у круглоротых. У некоторых хрящевых рыб (у ската Raja clavata) в каудальных отделах медианной зоны идентифицировано крупноклеточное нижнее ядро шва, которое проецируется в спинной мозг и участвует в иннервации электрического органа. В последние годы установлено, что клетки ядер шва костистых рыб серотонинергические. Медиальная зона ретикулярной формации рыб содержит нижнее, среднее и верхнее ретикулярные ядра. Они получают афференты от спинного мозга, тектума, ядер мозжечка. У костистых рыб в эту зону приходят проекции 5-10 пар черепномозговых нервов. Нисходящие (эфферентные) связи медиальной зоны приурочены главным образом к многокомпонентному медиальному продольному пучку, который соединяет ствол мозга с моторными отделами спинного мозга.
Рис. 13. Мозг акулы Galeus cams (по: Е.К. Сепп, 1959): 1 - обонятельная доля, 2 - обонятельный тракт, 3 - конечный мозг, 4 - зрительный нерв, 5 - крыша среднего мозга, 6 - блоковый нерв, 7 - мозжечок, 8 - тройничный нерв, 9 - ушки мозжечка, 10 - блуждающий нерв Наконец, третья - латеральная, зона ретикулярной формации состоит из мелких нейронов, которые образуют колонку. У некоторых акул, в пределах данной, зоны у места вхождения восьмого черепномозгового нерва описано «ядро В». Клетки, образующие это ядро, связаны с иннервацией вестибулолатеральной системы. Таким образом, ретикулярная формация рыб оказывается четко структурированной и содержит многие ядра, которые ранее считались характерными только для высших позвоночных. В ретикулярной формации осуществляется конвергенция различных сенсорных сигналов и имеются множественные выходы на моторные системы спинного мозга. Особенности строения ретикулярной формации позволяют рассматривать данную структуру как древнейшую интегративную систему мозга позвоночных животных. Важную роль в двигательной координации рыб играет мозжечок, который достигает значительной величины. Объем мозжечка увеличивается начиная с хрящевых рыб. Помимо ушек (аурикул), представляющих древний мозжечок, формируются корпус мозжечка и «заслонка», образующие старый мозжечок. Отделы мозжечка выполняют разные функции: первый из них (древний мозжечок) связан с вестибуло-латеральной системой, а второй (старый) включен в систему анализа проприоцептивной информации (рис. 13). Если у круглоротых мозжечок не дифференцирован по своей внутренней организации, то у рыб он представляет собой многослойную структуру, в которой можно выделить слои: молекулярный, клеток Пуркинье, фиброзный и зернистый. Хотя у рыб нет оформленных внутримозжечковых ядер, в белом веществе намечается скопление клеток, которое определяется как латеральное ядро. Часть нервных волокон, идущих от этого ядра, заканчивается в зачаточном красном ядре ретикулярной формации среднего мозга. Другая часть пучка направляется в гипоталамус и по пути отдает коллатерали к сетевидной формации заднего мозга и к ядрам глазодвигательных нервов. Афферентные связи мозжечка у рыб представлены спиномозжечковым, оливомозжечковым, гипоталамомозжечковым трактами, а также волокнами, идущими от крыши среднего мозга. Богатство связей мозжечка свидетельствует о том, что этот орган не только имеет большое значение в регуляции двигательной координации и тонуса туловищной мускулатуры, но и включен в систему восприятия и передачи импульсов от сенсорных систем. В частности, информация от зрительной системы может поступать в кору мозжечка через тектоцеребеллярный тракт от крыши среднего мозга. Это предположение, основанное на структурных предпосылках, было доказано Н.П. Веселкиным и К. Ковачевичем (1973) в условиях электрофизиологического эксперимента. Ими установлено, что у акул при стимуляции зрительного нерва или при электрическом раздражении крыши среднего мозга на поверхности тела мозжечка регистрируются вызванные потенциалы сложной конфигурации. Видимо, не случайно после удаления тела мозжечка у акул наблюдается выраженное нарушение сенсорной деятельности, в частности снижение остроты зрения, увеличение порога ответных реакций на раздражение органов боковой линии. Кроме того, послеоперационный синдром у акул проявляется в исчезновении выработанных пищевых условных рефлексов и в дезорганизации моторной деятельности: качание тела из стороны в сторону, нарушение движений хвоста. Все эти факты позволили А.И. Карамяну прийти к заключению, что мозжечок у рыб выступает одним из высших интегративных аппаратов, регулирующих моторные и сенсорные функции. Следующий важный координирующий центр головного мозга рыб - крыша среднего мозга. Так же как и мозжечок, она представляет собой многослойную структуру, построенную по экранному принципу. В тектуме акул описано до шести клеточных слоев: бедный клетками молекулярный, слой оптических волокон, слой главных клеток, внутренняя мозговая и внутренняя клеточная пластинки, перивентрикулярный слой, который содержит крупноклеточное ядро. У некоторых костистых рыб в коре тектума насчитывают 10 клеточных слоев, выраженных более четко, чем у пластиножаберных рыб. В тектуме веерообразно распространяются волокна зрительного тракта. Часть из них посылает коллатерали к наружному коленчатому телу промежуточного мозга. Кроме зрительных проекций в тектум у рыб приходят афферентные волокна от центров боковой линии и слуха, соматические афференты из длинного мозга. В то же время аксоны тектальных нейронов формируют обширные эфферентные связи с моторными ядрами спинного мозга, продолговатого мозга (тектобульбарные тракты), с гипоталамусом. Благодаря богатству связей крыша среднего мозга становится одним из центральных аппаратов, координирующих функции зрительной и соматосенсорной систем, принимающих участие в регуляции движений и реализации ряда жизненно важных безусловных рефлексов. К числу таких рефлексов можно отнести «сторожевой рефлекс», который вызывается раздражителями, предвещающими возможную опасность (надвиганием массивных предметов, резкими звуковыми и световыми раздражителями). Данный рефлекс проявляется в настораживании, приведении скелетной мускулатуры в состояние готовности. Вместе с тем кроме указанных выше стереотипных врожденных форм поведения средний мозг рыб участвует в реализации приобретенных форм поведения. В исследованиях А.В. Бару, В.Л. Бианки и др. (1960) было установлено, что условнорефлекторные реакции на световые раздражители могут быть выработаны у костистых рыб с удаленным передним мозгом. Если затем у экспериментальных животных удалить крышу среднего мозга, то условные рефлексы исчезают. Следовательно, крыша среднего мозга у рыб играет существенную роль в процессах сенсомоторной интеграции. У рыб, как и у более высокоорганизованных позвоночных, средний мозг участвует в регуляции движений и мышечного тонуса. При одностороннем удалении тектума у рыб происходит резкое возрастание тонуса сгибателей на стороне операции. В случае разобщения связей среднего мозга с продолговатым наблюдается повышенная двигательная активность плавников у рыб. Значит, от среднего мозга к исполнительным моторным центрам поступают тормозные влияния. Существенные структурные преобразования промежуточного мозга рыб, через который проходят пути афферентного снабжения конечного мозга, в значительной мере определяют дальнейший ход его эволюции. В самой дорсальной части промежуточного мозга - эпиталамусе, укрупняются ядра габенулярных ганглиев (ганглиев уздечки), к которым подходят пути от обонятельных областей конечного мозга. Расположенный вентральнее зрительный бугор (таламус), так же как и у круглоротых, разделен на дорсальную и вентральную части. В филогенетически более молодом образовании - дорсальном таламусе костистых рыб, можно вычленить основные группы ядер: медиальную (перивентрикулярную), интермедиальную и латеральную (поверхностную). Ядра: НКТ, круглое, дорсомедиальное переднее, дорсолатеральное переднее, вентромедиальное и др. более дифференцированы, чем у круглоротых и хрящевых рыб. С помощью морфологических методов в наружном коленчатом теле обнаружены проекции оптического тракта. Дорсального отдела таламуса достигают также волокна спиномезенцефального, бульбомезенцефального трактов, волокна, идущие из конечного мозга, крыши среднего мозга и мозжечка. Эфферентные пути представлены таламотеленцефальным и паллиальным трактами. Таким образом, уже на ранних этапах филогенеза таламус становится своеобразным коллектором афферентных путей, идущих к конечному мозгу - согласно классическим представлениям, надстройке над системой дистантных рецепторов. Гипоталамус рыб, так же как и таламус, претерпевает значительные изменения по сравнению с гипоталамусом круглоротых. У хрящевых рыб наблюдается более четкое деление гипоталамуса на передний, средний и задний отделы. Происходит дальнейшая дифференциация связей гипоталамуса с конечным мозгом, крышей среднего мозга и мозжечком. Вместе с тем резких границ между ядерными структурами гипоталамуса еще нет, и это в значительной мере затрудняет их идентификацию. Преоптическое ядро у рыб расположено в боковых стенках третьего желудочка. У костистых рыб нейросекреторные клетки этого ядра расположены довольно компактно. Ядро в целом обильно васкуляризировано, причем наблюдается тесный контакт капилляров с телами клеток. Аксоны нейросекреторных клеток преоптического ядра, формирующие парные мощные преоптико-гипофизарные пути, проходят через латеральное ядро серого бугра и далее воссоединяются в гипоталамо-гипофизарный тракт, идущий в нейрогипофиз. У костистых рыб различных видов нейрогипофиз имеет разное строение - от древовидного разветвленного до компактного образования. От тонких стволов нейрогипофиза в паренхиму аденогипофиза отходят концевые разветвления. Второй нейросекреторный центр гипоталамуса рыб представлен латеральным ядром серого бугра. Аксоны клеток этого ядра также направляются в нейрогипофиз. Гормон гипоталамо-гипофизарной нейро-секреторной системы рыб - вазотоцин играет существенную роль в регуляции водно-солевого обмена. Организация конечного мозга у хрящевых и костистых рыб неоднозначна по некоторым структурным показателям. По сравнению с круглоротыми у пластиножаберных рыб (акул, скатов) конечный мозг значительно увеличивается, особенно за счет развития его вентральной части. Мощные обонятельные доли (луковицы) выделяются из общей массы конечного мозга и соединяются с ним коротким обонятельным трактом (рис. 13). В обонятельной луковице имеется полость, она соединяется с боковым желудочком конечного мозга. Полушария конечного мозга отчетливо разъединяются только на ростральном конце, каудальнее стенки полушарий сливаются. От обонятельной луковицы проекционные волокна идут к латеральной и вентролатеральной поверхностям ипсилатерального полушария. Остальная часть конечного мозга (базальная, медиальная области) лишена прямых обонятельных проекций. При разрушении НКТ таламуса акул происходят дегенеративные изменения в аксонах таламотеленцефального тракта, по которому в конечный мозг, по-видимому, поступает информация от зрительных рецепторов. Эти экспериментальные факты свидетельствуют о том, что конечный мозг пластиножаберных рыб получает афференты различных модальностей, а не только выполняет чисто обонятельные функции. В пользу данной точки зрения говорят и данные электрофизиологических опытов. У ската при электрическом раздражении зрительного нерва в прилежащей к срединной линии обширной области паллиума конечного мозга регистрируются как суммарные вызванные потенциалы, так и импульсные ответы одиночных нейронов, синхронные с ритмом раздражения. В этой зоне можно отводить вызванные потенциалы при использовании соматических раздражителей (электрокожного раздражения). Наконец, в случае локального раздражения гипоталамуса от поверхности полушарий регистрируются медленные электрические колебания, многократно превышающие по амплитуде фоновую электрическую активность. Таким образом, в конечном мозге пластиножаберных рыб обнаружены проекции различных афферентных систем, что свидетельствует об участии этого отдела мозга в анализе афферентных сигналов. Примитивные черты организации проявляются, однако, в том, что проекции сенсорных систем в конечном мозге перекрываются и сохраняют элементы диффузности. По данным Н. Холмгрена (1925), в конечном мозге пластиножаберных нет четкого разграничения зачаточных кортикальных формаций, древней, старой и новой коры. Разделение плаща на латеральный (пириформный), общий, медиальный (гиппокампальный) паллиум обнаружено Холмгреном только у эмбрионов акулы Acanthias vulgaris. На более поздних стадиях развития (71 день) наблюдаются миграция клеток зачаточной общей коры в соседние зоны и стирание границ между отдельными кортикальными формациями. Однако позже в исследованиях Д.К. Обухова (1985) установлено, что конечный мозг колючей акулы отличается довольно высокой степенью нейронной дифференциации и содержит зачатки медиальной (гиппокампальной), дорсальной (общей) и латеральной (пириформной) коры. Наиболее сложная цитоархитектоника свойственна дорсальному паллиуму, состоящему из четырех слоев: наружного, пирамидного, внутреннего плексиморфного и эпендимы. Пирамидоподобные клетки второго слоя обладают развитой сетью апикальных и базальных дендритов. У костистых рыб выделить зачаточные кортикальные формации конечного мозга довольно трудно в связи с особым типом эмбриогенеза этого отдела ЦНС. В отличие от других позвоночных у костистых рыб конечный мозг развивается из парных дорсолатеральных выростов (боковых пузырей) ростральной части нервной трубки не путем эвагинации, а только вследствие эверсии (выворачивания) латеральной стенки мозга. Поэтому в конечном мозге костистых рыб нет боковых желудочков и полушария имеют вид сплошных долей, выстланных снаружи эпендимой. Данное обстоятельство затрудняет установление гомологии зачаточных кортикальных формаций у костистых рыб и у других позвоночных. Одни авторы, например А. Морган (1975), X. Шницлейн (1977), устанавливают гомологию на основе цитоархитектоники и распределения ферментов в различных структурах конечного мозга костистых рыб. Другие исследователи, в частности Р. Ньювенхус (1963), Р. Норткат (1995), Д. Обухов (1996), рассматривают конечный мозг костистых рыб как уникальную структуру и считают, что проведениегомологии с соответствующими кортикальными формациями других позвоночных крайне затруднено. Таким образом, данная проблема еще далека от своего разрешения. Из результатов физиологических экспериментов, которые проводятся параллельно с изучением структуры конечного мозга рыб, следует, что эта часть ЦНС играет существенную роль в организации врожденного, генетически детерминированного поведения. Так, например, у цихлидовых рыб после удаления конечного мозга наблюдается нарушение стайного поведения. Прооперированные рыбы образуют стаи значительно реже, чем интактные, причем стаи характеризуются малой прочностью. Одна из причин этого явления заключается в исчезновении разнообразных подражательных (имитационных) рефлексов, которые вносят значительный вклад в процесс стаеобразования. Удаление конечного мозга отрицательно сказывается на организации агрессивного и оборонительного поведения у костистых рыб. По данным И.В. Малюковой (1979), оперированные зеленушки (Grenttabrisgriseus) в море не занимают вновь своего охотничьего участка, утрачивают способность обороняться от вторжения конкурентов. У телапий (Telapia h. macrocephala) удаление конечного мозга приводит к снижению активности при градостроительстве и к нарушению способности самца оплодотворять икру. Вместе с тем функции конечного мозга рыб не ограничиваются регуляцией врожденных форм поведения. В равной мере удаление конечного мозга влияет на процессы обучения, ненаследственные поведенческие реакции. Хотя у костистых рыб с удаленным передним мозгом вырабатываются классические условные рефлексы, наблюдаются нарушение тонкой дифференциации условного сигнала, затруднения при решении сложных задач, которые содержат большое количество поведенческих ал |
Последнее изменение этой страницы: 2017-05-05; Просмотров: 515; Нарушение авторского права страницы