
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Энцефализация и процесс инвазии у круглоротых
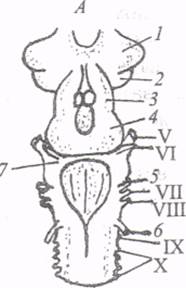
Исследование нервной системы круглоротых, находящихся филогенетически между бесчерепными и рыбами, представляет значительный интерес для эволюционной нейрофизиологии. Этот интерес обусловлен тем, что у круглоротых впервые в ряду древних позвоночных формируется головной мозг и практически все основные отделы ЦНС представлены как самостоятельные структурные образования. Спинной мозг миноги имеет вид плоской ленты с развитыми боковыми частями. Его внутренняя организация сохраняет черты примитивизма, свойственные бесчерепным. К таким особенностям относятся: раздельность чувствительных и двигательных нервов, отсутствие миелиновой оболочки на нервных волокнах, диффузность серого вещества, которое еще не разделяется на передние и задние рога. Вместе с тем в отличие от ланцетника у миноги часть нейронов, обеспечивающих кожную чувствительность, выделяется из спинного мозга и концентрируется в спинномозговых ганглиях. Наблюдается совершенствование проводниковой системы спинного мозга. Восходящие пути представлены филогенетически древней системой латеральных столбов, волокна которых оканчиваются главным образом в ретикулярной формации продолговатого мозга. Восходящие пути дорсальных столбов состоят из коротких звеньев, ориентированных в ростральном направлении. Они достигают продолговатого мозга. Нисходящие пути, соединяющие головной мозг со спинным, формируются у миноги ретикулоспинальными волокнами и системой гигантских (мюллеровских) волокон, расположенных вентральнее спинномозгового канала, аксонов клеток, лежащих в области продолговатого и среднего мозга. Таким образом, формирование головного мозга, или энцефализация, сопряжено с процессом инвазии, т. е. внедрением новых путей в структуры, ранее между собой не связанные. Как же можно себе представить процесс энцефализации? Согласно классическим представлениям Ч. Шеррингтона, А.А. Заварзина, Дж. Бишопа, головной мозг возник в связи с переходом животных к билатеральной симметрии тела и с концентрацией экстерорецепторов на переднем конце тела, как своеобразная надстройка над системами дистантной рецепции. Развитие вестибуло-латеральной системы, возникновение системы блуждающего нерва и дыхательного центра создало базу для формирования заднего мозга. Толчком к закладке крыши среднего мозга (tectum opticum) послужило развитие органов зрения; наконец, становление обонятельной рецепции, согласно классическим представлениям, стало стимулом для развития конечного мозга. Хотя филогенетическое становление различных отделов мозга могло происходить параллельно, однако в общем плане задний мозг явился надстройкой над спинным, средний над задним, а передний над всей предыдущей системой. На всех трех этапах надстройки головного мозга над спинным происходило подтягивание проводящих путей из нижележащих отделов к новым формациям. Далее А. Капперс, С.Д. Геррик, проведя фундаментальные исследования, высказали гипотезу о линейном увеличении относительных размеров головного мозга в ходе эволюции и о перемещении интегративных функций в ростральные, филогенетически наиболее молодые отделы мозга. Эти положения долгое время воспринимались как хрестоматийные. Однако после достижений сравнительной нейрологии последних лет стало ясно, что они не выдерживают критики. Так, например, линейного увеличения относительных размеров головного мозга (прежде всего его переднего отдела) в ряду позвоночных не происходит. Правильнее говорить о независимых процессах развития различных отделов мозга на определенных этапах филогенеза. В качестве иллюстрации можно привести увеличение размеров мозжечка у хрящевых рыб. В пределах отдельных классов позвоночных эволюция координационных систем мозга шла нередко параллельно, и это ставит под вопрос незыблемость принципа о каудоростральной последовательности возникновения отделов мозга. Окончательное решение этой проблемы в значительной степени осложняется также и тем, что среди нейроморфологов нет единодушия в суждениях о гомологии идентичных отделов мозга (например, таламических ядер) у позвоночных разных классов. Испытание временем не выдержала также концепция о надстраивании переднего мозга над обонятельной системой. Оказалось, что даже у низших позвоночных (круглоротых) конечный мозг неверно считать чисто обонятельным. В нем обнаружены проекции и необонятельных сенсорных систем - зрительной, соматической, которые вносят существенный вклад в формирование структуры конечного мозга. Установлено, что конечный мозг низших позвоночных участвует в реализации наиболее пластичных приобретенных форм поведения. В связи с этим определенной ревизии подвергается также представление о перемещении интегративных функций в ростральном направлении от каудальных, более древних нервных центров к ростральным, филогенетически более молодым структурам мозга. Термин «перемещение» не вполне адекватно отражает ситуацию, так как древние, каудальные сенсорные центры (например, среднего мозга) сохраняют свои базисные функции, как у низших, так и у высших позвоночных. Суть проблемы состоит не в перемещении в конечный мозг функций каудальных центров в плане их дублирования. Правильнее говорить о возникновении новых уровней интеграции в ростральных отделах мозга, о поэтапной функциональной надстройке. Процессы дифференциации и специализации в филогенетически наиболее молодых отделах мозга позволяют им осуществлять более детальный анализ информации, поступающей из окружающего мира. Это свойство отличает ростральные отделы от более жестко запрограммированных каудальных и позволяет новым формациям приобрести доминирующее положение в иерархии нервных центров. В целом вся система работает на основе функционального единства старых и новых отделов мозга. Исходя из общих установок попытаемся проследить, каким образом некоторые из этих принципов реализуются в конструкции головного мозга круглоротых. Общий план конструкции мозга миноги сходен с таковым у более высокоорганизованных позвоночных. Он состоит из четырех отделов: конечного мозга, промежуточного, среднего и ромбовидного вместе с мозжечком. Формируясь в ходе эмбриогенеза из третьего мозгового пузыря, ромбовидный мозг дифференцируется на продолговатый мозг и слабо развитый мозжечок в виде боковых выпячиваний нервной трубки (рис. 12). Полость ромбенцефалона превращается в четвертый желудочек мозга. От продолговатого мозга отходят корешки 5-10-й пары черепномозговых нервов, в том числе блуждающего (10-го), вестибуло-кохлеарного (8-го), лицевого (7-го) и тройничного (5-го). В передней половине продолговатого мозга локализованы ядра вестибуло-латеральной системы, куда приходят импульсы от рецепторов боковой линии, двух полукружных каналов и статоциста. Здесь, на уровне продолговатого мозга замыкаются рефлексы, направленные на поддержание правильного положения тела в пространстве и ориентацию. В толще продолговатого мозга расположена сетчатая, или ретикулярная, формация ствола, Рис. 12. Головной мозг миноги (по: А.И. Карамян, 1976): А - схематическое изображение с дорсальной поверхности, Б - фронтальный срез через конечный мозг: 1 - обонятельная луковица, 2 - конечный мозг, 3 - промежуточный мозг, 4 - крыша среднего мозга, 5, 6 - передний и задний латеральные нервы, 7 - мозжечок, 8 - гиппокампальный паллиум, 9 - дорсальный участок плаща, 10 - латеральный пириформный паллиум, 11 - полосатое тело, 12 - обонятельная луковица, 13 - переднее, дорсальное ольфакторное ядро; V-X - черепномозговые нервы Мозжечок у миноги является своеобразной надстройкой над ядрами органа боковой линии и вестибуло-латеральной системы. Несмотря на примитивную организацию мозжечок миноги имеет довольно богатые эфферентные связи с ядрами продолговатого мозга и покрышкой среднего мозга, В белом веществе мозжечка идентифицированы аксоны клеток, которые образуют пучок - предшественник ножек мозжечка. По данным некоторых авторов, в сером веществе мозжечка миноги имеются два типа клеток: мелкие зернистые и более крупные, напоминающие по своим очертаниям клетки Пуркинье. В составе среднего мозга миноги выделяют тектальную область (tectum), или крышу среднего мозга, - гомолог переднего двухолмия и расположенную вентральнее - покрышку (tegmentum), где локализован torus semicircularis - зачаток заднего двухолмия. В крышу среднего мозга проецируются волокна зрительного тракта, восходящие волокна вестибуло-латеральной системы и пути, передающие соматическую информацию от спинного мозга. От глубоких слоев тектума начинаются эфферентные пути, связывающие крышу среднего мозга с моторными центрами продолговатого и спинного мозга, с мозжечком и промежуточным мозгом. Таким образом, структура крыши среднего мозга создает предпосылки для ее участия в анализе сенсорной информации и регуляции движений. Разряды эфферентных нейронов тектума реализуются в форме ряда жизненно важных, стереотипных безусловных рефлексов. Зона torus stmicircularis представляет собой закладку будущих задних бугров четверохолмия, которые полностью формируются у рептилий. У миноги в эту зону приходят пути от вестибуло-латеральной системы продолговатого мозга. Тегментум содержит ядра глазодвигательного и блокового нервов. В дорсальном и вентральном отделах тегментума локализованы тела трех пар гигантских клеток Мюллера, аксоны которых вместе с отростками нейронов ретикулярного ядра присоединяются к медиальному продольному пучку и идут в продолговатый и спинной мозг. Двусторонними трактами тегментум связан и с вышележащими отделами мозга, в частности с промежуточным мозгом. У круглоротых промежуточный мозг подразделяется на три основных отдела: эпиталамус, таламус и гипоталамус. Эпиталамус (надбугорье) представлен парным габенулярным ганглием, связанным с развитием теменного органа и получающим проекции от обонятельной системы. Таламус (зрительный бугор) миноги слабо дифференцирован, его ядра представляют собой специализированные участки нейропиля, изолированные от клеточных тел. Вместе с тем в дорсальном таламусе миноги можно выделить дорсальное ядро и зачаточное наружное коленчатое тело (НКТ), куда приходят зрительные афференты. Клетки НКТ различны по величине, их дендриты распространяются среди длинных пучков, а аксоны присоединяются к нескольким восходящим и нисходящим трактам, в том числе к таламофронтальному, который достигает конечного мозга. Этот тракт служит важным источником афферентного снабжения конечного мозга, тем каналом, по которому поступает информация от зрительного тракта, вестибуло-латерального лемниска. Волокна этих трактов образуют проекции на клетках НКТ, которое таким образом становится промежуточной «станцией» в системе афферентного снабжения конечного мозга. В вентральной части таламуса миноги идентифицированы вентральное ядро и его продолжение - ядро заднего бугорка. Эта область промежуточного мозга имеет связи с конечным мозгом, гипоталамусом и с моторной зоной покрышки среднего мозга, участвуя, следовательно, в моторной координации. Гипоталамус (подбугорье) миног имеет план строения, сходный с таковым у более высокоорганизованных позвоночных. Гипоталамус состоит из дорсальной и вентральной частей, причем последняя разделяется на переднюю и заднюю доли, ядра постоптической, преинфундибулярной, инфундибулярной комиссур и воронку (infundibulum). Ядра гипоталамуса имеют связи с таламусом, с обонятельной системой конечного мозга, базальными ганглиями. Таким образом, гипоталамус, так же как и таламус, является одним из источников афферентного снабжения конечного мозга. В гипоталамусе миноги, в непосредственной близости от полости третьего желудочка расположено крупноклеточное преоптическое ядро. Оно содержит биполярные нейросекреторные клетки, аксоны которых формируют идущий к гипофизу преоптико-гипофизарный тракт. Сформированного нейрогипофиза у круглоротых нет, его гомологом выступает нейрогипофизарный слой нервных волокон, который лежит между вентральной стенкой воронки и дорсальной поверхностью аденогипофиза. Значит, можно говорить о том, что у круглоротых формируются все основные компоненты гипоталамо-гипофизарной нейросекреторной системы. Конечный мозг круглоротых характеризуется примитивной внутренней организацией, в частности диффузной, нейропильной структурой и перивентрикулярным положением основной массы клеток, из которых только часть мигрировала в наружный слой. Дифференцировка на паллиальные (плащевые) и ядерные (полосатое тело) структуры только намечается, и они мало отличаются по своей организации. Между различными формациями переднего мозга нет четких границ, что создает значительные трудности при идентификации тех илииных образований. Тем не менее, в конечном мозге круглоротых можно выделить несколько зачаточных образований плаща, которые на последующих этапах эволюции развиваются в три формации коры (древнюю, старую и новую), имеющие свои специфические источники афферентного снабжения. Конечный мозг миноги состоит из так называемых парных полушарий и срединной неразделенной части (рис. 12). В состав полушария входит обонятельная луковица, в которую дорсомедиально внедряется обонятельный нерв. Волокна этого нерва образуют поверхностный слой луковицы. Несколько глубже находится клубочковый слой, в котором локализованы крупные обонятельные гломерулы, окруженные митральными клетками. По структуре обонятельные луковицы достаточно консервативны и сохраняют основной принцип организации у всех позвоночных, а их развитие идет по пути дифференциации клеточных слоев и их связей. Вентромедиально луковица, переходит в обонятельную долю полушария. От обонятельной луковицы в полушарие тянется обонятельный тракт, волокна которого проецируются в латеральный пириформный паллиум, представляющий собой зачаток древней (paleocortex) обонятельной коры (рис. 12). Волокна обонятельного тракта частично достигают гиппокампального паллиума, частично полосатого тела - стриатума. Зачаток обонятельной коры - латеральный пириформный паллиум расположен латеро-вентрально по отношению к заднему рогу бокового желудочка и граничит с передним ольфакторным ядром, стриатумом, а также с дорсальным участком плаща (рис. 12). Пириформный паллиум, как и весь плащ (паллиум), формирующий крышу переднего мозга, характеризуется перивентрикулярным положением основной массы клеток. Они расположены группами и посылают свои отростки в сторону стриатума и в переднее ольфакторное ядро. Пириформный паллиум пересекается волокнами ольфакторного тракта и вентрального ольфакто-таламического и гипоталамического тракта. Дорсомедиальная неэвагинированная часть конечного мозга миноги занята гиппокампальным паллиумом, зачатком старой, гиппокампальной коры (archicortex) (рис.12). Д. Джонстон (1902) впервые определил гиппокампальный паллиум (ГП) как гомолог гиппокампа высших позвоночных. Эта точка зрения имеет последователей и противников, которые относят ГП миноги к промежуточному мозгу. У миноги ГП отличается от других паллиальных образований тонкой структурой и богатством связей. Это проявляется в миграции нейронов и в их дифференцированности. Нейроны грушевидной или пирамидной формы располагаются в нескольких слоях. Толстые дендриты этих клеток усеяны шипиками, что является отличительной особенностью нейронов ГП. О.А. Поленовой и Н.П. Веселкиным (1988) с помощью пероксидазы хрена впервые выявлены прямые таламо-гиппокампальные связи, которые рассматриваются как субстрат поступления необонятельной (зрительной, акустической, соматической) информации в конечный мозг миноги. Висцеральная информация в ГП поступает из гипоталамуса. Обнаружены также двусторонние связи ГП миноги с обонятельными луковицами. Таким образом, ГП миноги обладает двойной афферентацией: экстероцептивной и висцеральной. Наконец, третья зачаточная корковая формация конечного мозга миноги представлена дорсальными участками плаща (рис. 12), расположенными между пириформным и гиппокампальным паллиумом. Дорсальный участок плаща - это предшественник зачатка новой коры, появляющейся у более высокоорганизованных позвоночных животных. Вопрос о функциях конечного мозга круглоротых детально разрабатывался в лаборатории А.И. Карамяна. Электрофизиологическими методами Н.П. Веселкин, А.Г. Агаян. П. Брукмозер и др. (1963-1973) установили распределение необонятельных сенсорных проекций в головном мозге миног. В случае электрической стимуляции зрительного нерва его проекции обнаружены в гиппокампальном паллиуме. Эти проекции перекрываются с проекциями обонятельных афферентов. Здесь же, в ГП регистрировались вызванные потенциалы в ответ на раздражение дорсальной поверхности спинного мозга, где проходят соматические афференты. Анализ указанных выше экспериментальных фактов позволил А.И. Карамяну и Н.П. Веселкину сделать вывод о том, что конечный мозг круглоротых нельзя рассматривать как чисто обонятельный, так как в нем обнаружены проекции необонятельных систем (зрительной и соматической). Второе важное заключение сводилось к тому, что необонятельные сенсорные системы круглоротых имеют переключение в таламических ядрах. В этом состоит принципиальное сходство в организации центральных отделов сенсорных систем у домлекопитающих позвоночных и млекопитающих. Очевидно, три зачаточные формации конечного мозга имеют три источника афферентного снабжения: от обонятельной системы, таламуса и гипоталамуса. Все эти источники играют существенную роль в ходе дальнейшего прогрессивного развития конечного мозга. Необонятельные проекции в гиппокампальном паллиуме (зачаток гиппокампальной коры) свидетельствуют о том, что здесь осуществляется конвергенция информации, поступающей как от экстеро-, так и от интерорецепторов. На базе этих процессов осуществляется интеграция соматических и вегетативных реакций в целостные поведенческие акты. Архипаллиум (ГП) круглоротых сравним с гиппокамгюм в лимбической системе высших позвоночных, которая ответственна за осуществление пищевого, полового, оборонительного поведения. Лимбическая система имеет такое же древнее: происхождение, как и обонятельная, и, естественно, вносит существенный вклад в реализацию интегративной функции конечного мозга. Исследовать поведенческие акты миног в лабораторных условиях сложно, так как эти животные малоподвижны в неволе. Вместе с тем в экспериментах А.В. Бару, В.В. Фанарджяна, Б.Ф. Сергеева (1967) была выявлена возможность образования условных реакций у миног на световые и вибрационные раздражители. Эти приобретенные оборонительные реакции квалифицируются как условные по целому ряду показателей. Во-первых, они возникают только при сочетании условного и безусловного (электрический ток) раздражителей. Во-вторых, выработанные реакции характеризуются относительной стабильностью и сохраняются в течение нескольких дней. Наконец, в-третьих, условные оборонительные реакции миног могут быть экстренно угашены и вновь восстановлены после нескольких подкреплений условного раздражителя. Однако приобретенным реакциям круглоротых свойственны и черты примитивизма, которые сближают их с суммационными рефлексами бесчерепных. У миног возможна выработка оборонительных условных реакций на одномодальные раздражители различной силы. Так же как у ланцетника, у миноги из всех форм внутреннего торможения вырабатывается только угасательное. Низкий уровень высшей нервной деятельности круглоротых проявляется и в отсутствии анализа комплексных раздражителей, и в невозможности выработки условных рефлексов высшего порядка, образованных на базе закрепленного условного рефлекса первого порядка. Учитывая «двойственность» приобретенных реакций круглоротых, которые, с одной стороны, имеют некоторое сходство с условными рефлексами, а с другой - сохраняют особенности суммационных рефлексов, А. И. Карамян выделил эти реакции в промежуточную группу явлений и определил как квазиусловные рефлексы. Следует еще раз отметить, что у круглоротых формируется головной мозг, общая схема конструкции которого идентична плану организации мозга более высокоорганизованных позвоночных. Мозг как надсегментарная система интеграции участвует в реализации стереотипных безусловных рефлексов и в программировании более сложных поведенческих актов, сочетающих соматические и вегетативные компоненты реакций. В качестве аппарата координации нервной деятельности у круглоротых используются не только стволовые структуры мозга (продолговатый, средний мозг), но и конечный мозг, где в зачаточном состоянии закладываются три формации коры головного мозга. Конечный мозг круглоротых не является чисто обонятельным и получает экстероцептивную информацию, которая используется в процессе сенсомоторной интеграции. Вместе с тем в ЦНС круглоротых сохраняется много черт диффузной организации. Эти черты проявляются и в строении серого вещества спинного мозга, и в слабой дифференциации таламических ядер, и в нейропильной структуре конечного мозга, где большинство клеток занимает перивентрикулярное положение. Перекрытие проекций обонятельных, зрительных и соматических афферентов в конечном мозге и стволовых образованиях миноги также свидетельствует о диффузной организации ЦНС. Несовершенство структурной специализации мозга сопряжено с примитивизмом приобретенных реакций круглоротых, которые можно отнести к квазиусловным рефлексам. Каким же образом совершался дальнейший переход от диффузности к специализации? На этот вопрос мы найдем ответ при знакомстве со структурой и функцией ЦНС позвоночных, стоящих на более высоких ступенях эволюционного развития. Ихтиопсидный этап развития Морфофизиологические преобразования в спинном мозге рыб сопряжены с появлением более сложных форм двигательного поведения. В частности, переход от стереотипных ундулирующих движений круглоротых к локомоции с помощью плавников связан с дальнейшей дифференциацией кожной и мышечно-суставной (проприоцептивной) чувствительности. У хрящевых пластиножаберных рыб в спинальных ганглиях имеются специализированные нейроны для выполнения этих функций. На базе проприоцептивной чувствительности совершенствуются физиологические механизмы рефлекторного сопротивления мышцы ее растяжению, механизмы динамической фиксации суставов. Наблюдается постепенная концентрация серого вещества в окружности центрального канала. Укорачивается путь моторных аксонов внутри спинного мозга, причем каждый мотонейрон иннервирует не миомер (как у круглоротых), а определенную группу мышц плавника. Белое вещество спинного мозга рыб также претерпевает определенную дифференциацию. Нисходящие пути представлены древним ретикулоспинальным трактом. Среди нейронов, образующих этот тракт, особо выделяются маутнеровские клетки, лежащие в продолговатом мозге Гигантские аксоны маутнеровских клеток посылают множество коллатералей к вставочным и моторным нейронам спинного мозга. Раздражение ретикуло-спинального тракта вызывает ипсилатерально сокращения мышц туловища, жаберной крышки, нижней челюсти; в то же время на противоположной стороне туловища наблюдается торможение моторики. У хрящевых пластиножаберных рыб появляются в виде особых пучков вестибуло-спинальный и церебелло-спинальный тракты, которые связывают спинной мозг с центральными аппаратами регуляции движения (вестибулярным аппаратом и мозжечком). Эти связи двусторонние и дополняются восходящим спино-церебеллярным трактом, проходящим в латеральных столбах от уровня шейной часта: спинного мозга. Спиноцеребеллярный тракт рыб достигает корпуса мозжечка и может быть отождествлен с дорсальным мозжечковым трактом Флексига высших позвоночных. Кроме того, группа восходящих волокон вентролатеральных столбов оканчивается в ретикулярной формации продолговатого мозга и частично в крыше среднего мозга. Сравнительно недавно окончания восходящих спинальных волокон обнаружены у акул в таламусе промежуточного мозга. Долго считали, что филогенетически более молодые восходящие пути дорсальных столбов у рыб, как и у миног, представляют внутреннюю эндоспинальную систему и не имеют проекций в стволовых частях мозга. Однако недавно в дорсальных столбах хрящевых и костистых рыб идентифицированы длинные спинальные волокна в области спинобульбарного соединения. Электрофизиологическими методами доказано участие дорсальных столбов у костистых рыб, как и у круглоротых, в проведении импульсации, вызванной тактильным раздражением. Таким образом, в процессе филогенетического развития происходит постепенное расширение связей спинного мозга с различными отделами головного мозга. Непосредственное продолжение спинного мозга - продолговатый мозг у рыб имеет ряд продольных возвышений, содержащих ядра черепномозговых нервов. Среди них следует отметить ядра вестибулолатеральной системы, с помощью которой осуществляется рефлекторная установка головы и тела в пространстве. Дифференциация рецепторов боковой линии, появление у хрящевых рыб третьего полукружного канала в лабиринте сопряжено со значительным увеличением объема ядер вестибуло-латеральной системы. Дорсальное ядро, в котором оканчиваются толстые волокна переднего нерва боковой линии, превращается в целую долю. Одновременно разрастается медиальное ядро, к которому подходят тонкие волокна переднего и заднего нервов боковой линии. В вентральном ядре, куда проецируются окончания восьмого нерва (вестибулокохлеарного), увеличивается число составляющих это ядро клеток, они становятся крупнее. Слуховые функции у рыб выполняют нижняя часть лабиринта внутреннего уха (саккулюс) и в качестве усиливающего воспринимающего прибора плавательный пузырь. Отметим, что пластино-жаберные рыбы перестают реагировать на низкочастотные звуки (500-600 Гц) после перерезки восьмого нерва. Соматическая афферентная зона в продолговатом мозге рыб представлена помимо системы вестибулокохлеарного нерва системой тройничного нерва (пятая пара). Восходящие проекции ядер тройничного нерва направляются к среднему (тектуму, тегментуму), промежуточному и конечному мозгу. Эфференты ядер тройничного нерва адресованы моторным ядрам стволовой части мозга и ретикулярной формации. У акул эфференты ядер тройничного нерва идут в спинной мозг, часть входов направляется в мозжечок. Чувствительные окончания тройничного нерва рыб имеются в слизистой оболочке ротовой полости. Их возбуждение при наполнении полости рта водой активирует инспираторные нейроны дыхательного центра, осуществляющего автоматическую смену фаз дыхательного цикла. В рефлекторной регуляции дыхательных движений принимают участие также афферентные волокна лицевого (седьмая пара) и блуждающего (десятая пара) нервов. Ядра этих нервов формируют висцеросенсорную зону продолговатого мозга. Сюда поступает информация от хеморецепторов, сосредоточенных на поверхности головы, в жабрах, в глотке. Данная система обусловливает восприятие природы химических и пищевых компонентов окружающей среды. Нейроны моторной части ядра блуждающего нерва координируют деятельность пищеварительного тракта и других внутренних органов. В сетчатой (ретикулярной) формации продолговатого мозга рыб выделяют медианную, медиальную и латеральную зоны. В медианной зоне локализованы переднее, среднее и заднее ядра шва, которые описаны у круглоротых. У некоторых хрящевых рыб (у ската Raja clavata) в каудальных отделах медианной зоны идентифицировано крупноклеточное нижнее ядро шва, которое проецируется в спинной мозг и участвует в иннервации электрического органа. В последние годы установлено, что клетки ядер шва костистых рыб серотонинергические. Медиальная зона ретикулярной формации рыб содержит нижнее, среднее и верхнее ретикулярные ядра. Они получают афференты от спинного мозга, тектума, ядер мозжечка. У костистых рыб в эту зону приходят проекции 5-10 пар черепномозговых нервов. Нисходящие (эфферентные) связи медиальной зоны приурочены главным образом к многокомпонентному медиальному продольному пучку, который соединяет ствол мозга с моторными отделами спинного мозга.
Рис. 13. Мозг акулы Galeus cams (по: Е.К. Сепп, 1959): 1 - обонятельная доля, 2 - обонятельный тракт, 3 - конечный мозг, 4 - зрительный нерв, 5 - крыша среднего мозга, 6 - блоковый нерв, 7 - мозжечок, 8 - тройничный нерв, 9 - ушки мозжечка, 10 - блуждающий нерв Наконец, третья - латеральная, зона ретикулярной формации состоит из мелких нейронов, которые образуют колонку. У некоторых акул, в пределах данной, зоны у места вхождения восьмого черепномозгового нерва описано «ядро В». Клетки, образующие это ядро, связаны с иннервацией вестибулолатеральной системы. Таким образом, ретикулярная формация рыб оказывается четко структурированной и содержит многие ядра, которые ранее считались характерными только для высших позвоночных. В ретикулярной формации осуществляется конвергенция различных сенсорных сигналов и имеются множественные выходы на моторные системы спинного мозга. Особенности строения ретикулярной формации позволяют рассматривать данную структуру как древнейшую интегративную систему мозга позвоночных животных. Важную роль в двигательной координации рыб играет мозжечок, который достигает значительной величины. Объем мозжечка увеличивается начиная с хрящевых рыб. Помимо ушек (аурикул), представляющих древний мозжечок, формируются корпус мозжечка и «заслонка», образующие старый мозжечок. Отделы мозжечка выполняют разные функции: первый из них (древний мозжечок) связан с вестибуло-латеральной системой, а второй (старый) включен в систему анализа проприоцептивной информации (рис. 13). Если у круглоротых мозжечок не дифференцирован по своей внутренней организации, то у рыб он представляет собой многослойную структуру, в которой можно выделить слои: молекулярный, клеток Пуркинье, фиброзный и зернистый. Хотя у рыб нет оформленных внутримозжечковых ядер, в белом веществе намечается скопление клеток, которое определяется как латеральное ядро. Часть нервных волокон, идущих от этого ядра, заканчивается в зачаточном красном ядре ретикулярной формации среднего мозга. Другая часть пучка направляется в гипоталамус и по пути отдает коллатерали к сетевидной формации заднего мозга и к ядрам глазодвигательных нервов. Афферентные связи мозжечка у рыб представлены спиномозжечковым, оливомозжечковым, гипоталамомозжечковым трактами, а также волокнами, идущими от крыши среднего мозга. Богатство связей мозжечка свидетельствует о том, что этот орган не только имеет большое значение в регуляции двигательной координации и тонуса туловищной мускулатуры, но и включен в систему восприятия и передачи импульсов от сенсорных систем. В частности, информация от зрительной системы может поступать в кору мозжечка через тектоцеребеллярный тракт от крыши среднего мозга. Это предположение, основанное на структурных предпосылках, было доказано Н.П. Веселкиным и К. Ковачевичем (1973) в условиях электрофизиологического эксперимента. Ими установлено, что у акул при стимуляции зрительного нерва или при электрическом раздражении крыши среднего мозга на поверхности тела мозжечка регистрируются вызванные потенциалы сложной конфигурации. Видимо, не случайно после удаления тела мозжечка у акул наблюдается выраженное нарушение сенсорной деятельности, в частности снижение остроты зрения, увеличение порога ответных реакций на раздражение органов боковой линии. Кроме того, послеоперационный синдром у акул проявляется в исчезновении выработанных пищевых условных рефлексов и в дезорганизации моторной деятельности: качание тела из стороны в сторону, нарушение движений хвоста. Все эти факты позволили А.И. Карамяну прийти к заключению, что мозжечок у рыб выступает одним из высших интегративных аппаратов, регулирующих моторные и сенсорные функции. Следующий важный координирующий центр головного мозга рыб - крыша среднего мозга. Так же как и мозжечок, она представляет собой многослойную структуру, построенную по экранному принципу. В тектуме акул описано до шести клеточных слоев: бедный клетками молекулярный, слой оптических волокон, слой главных клеток, внутренняя мозговая и внутренняя клеточная пластинки, перивентрикулярный слой, который содержит крупноклеточное ядро. У некоторых костистых рыб в коре тектума насчитывают 10 клеточных слоев, выраженных более четко, чем у пластиножаберных рыб. В тектуме веерообразно распространяются волокна зрительного тракта. Часть из них посылает коллатерали к наружному коленчатому телу промежуточного мозга. Кроме зрительных проекций в тектум у рыб приходят афферентные волокна от центров боковой линии и слуха, соматические афференты из длинного мозга. В то же время аксоны тектальных нейронов формируют обширные эфферентные связи с моторными ядрами спинного мозга, продолговатого мозга (тектобульбарные тракты), с гипоталамусом. Благодаря богатству связей крыша среднего мозга становится одним из центральных аппаратов, координирующих функции зрительной и соматосенсорной систем, принимающих участие в регуляции движений и реализации ряда жизненно важных безусловных рефлексов. К числу таких рефлексов можно отнести «сторожевой рефлекс», который вызывается раздражителями, предвещающими возможную опасность (надвиганием массивных предметов, резкими звуковыми и световыми раздражителями). Данный рефлекс проявляется в настораживании, приведении скелетной мускулатуры в состояние готовности. Вместе с тем кроме указанных выше стереотипных врожденных форм поведения средний мозг рыб участвует в реализации приобретенных форм поведения. В исследованиях А.В. Бару, В.Л. Бианки и др. (1960) было установлено, что условнорефлекторные реакции на световые раздражители могут быть выработаны у костистых рыб с удаленным передним мозгом. Если затем у экспериментальных животных удалить крышу среднего мозга, то условные рефлексы исчезают. Следовательно, крыша среднего мозга у рыб играет существенную роль в процессах сенсомоторной интеграции. У рыб, как и у более высокоорганизованных позвоночных, средний мозг участвует в регуляции движений и мышечного тонуса. При одностороннем удалении тектума у рыб происходит резкое возрастание тонуса сгибателей на стороне операции. В случае разобщения связей среднего мозга с продолговатым наблюдается повышенная двигательная активность плавников у рыб. Значит, от среднего мозга к исполнительным моторным центрам поступают тормозные влияния. |
Последнее изменение этой страницы: 2017-05-05; Просмотров: 517; Нарушение авторского права страницы

 выполняющая функции интеграции и связи со спинным мозгом.
выполняющая функции интеграции и связи со спинным мозгом.