|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
РАЗОБЩЕНИЕ ОХ И ФОСФОРИЛ.
Разобщители – в-ва, кот. разрешают перенос е, но не дают возм. синт. АТР → обр. ΔμН+, но эн. не идет в АТР. Принцип действия разобщителей: обеспеч. утечку Н+ через внутр. мембр., сами переносят Н+ или заставляют сделать это др. в-ва → сниж. ур-нь ΔμН+<V. Разобщители: экзо-/эндогенные. Эндогенные: (1) в-ва высшей физиологич. нормы (билирубин, тироксин, ненас. ВЖК, фенолы). (2) спец. белки-разобщ. (биол. роль – адаптация орг-ма к пониж. тем-рам). Экзогенные (ксенобиотки): (1) хим. в-ва, явл. ионофорами: (1а) хим. соед.: 2,4-динитрофенол, динитрокрезол, пентахлорфенол. (1б) антибиотики: грамицидин, валиномицин. (1в) антикоагулянты непрямого действия (трансп. Са2+). След. пирогенный эффект (экзо- 1б, 1в; эндо- 1). (2) в-ва, повр. мембраны: детергенты, АФК.
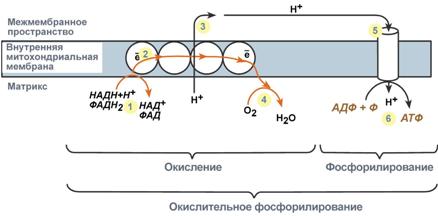
(31) Дыхательная цепь и теплопродукция. Коэффициент Р/О. Потоки важнейших метаболитов, поступающих в митохондрии и выходящих из них. Строение подробно – см. 65. Принцип работы дыхательной цепи 7. Образ. в р-иях катаб. NADH и FADH2 передают атомы водорода (т.е. H+ и e) на ферм. дых.цепи. 8. e движ. по ферментам дых.цепи и теряют энергию. 9. Эта энергия используется на выкач. Н+ из матрикса в межмембр. пр-во. 10. В конце дых.цепи e попадают на O2 и восст. его до H2O. 11. Н+ стремятся обратно в матрикс и проходят через АТФ-синтазу. 12. При этом они теряют энергию, которая исп. для синтеза ATP.
Т.о., восст. формы NADH+H+ и FADH2 окисляются → присоединение фосфата к ADP, т.е. фосфорилирование. Поэтому все вместе = окислительное фосфорилирование. СТРОЕНИЕ МТХ: внеш. мембр. проницаема для в-в с М 5-10 кДа (связ. с бол. кол-вом порина, форм. отв. d2-3нм; белок:липид=1:1), внутр. мембр. низкопроницаема, обр. кристы. Легко диффундируют только газы и мол. с М до 150 кДа. До 70% - белки, среди липидов – кардиолипин. Оч. гидрофобна, непрониц. для ионов. Для переноса разл. в-в организованы транслоказы, кот. переносят ADP, ПВК в матрикс, АТР, цитрат, Ас – из матрикса.
(32) Системы микросомального окисления. Строение, изоформы цитохрома Р450. Участие в эндогенном обмене и детоксикации. Микрос. Ох лок. в мембр. sER. Катализ. Ох больш-ва эндогенных субстр. и ксенобиотиков. Исп. мол. О2 и NADPH+H.
Гидроксилирующая сис-ма сост. из 2 комп.: (1) цит.Р450-редуктаза (сод. FAD, FMN). забир. е и Н+ с NADPH+H+, передает на субстрат. (2) цит.Р450 связ. одновр. О2 и субстрат, осущ. гидроксилир. Гемопротеин. 450 – max поглощения в комплексе с СО. Одноврем. связ. и субстрат, и О2.
Цит.Р450 найден ао всех ядросод. кл., оч. много в микросомах печени, в над/поч. Ф-ии микрос. Ох: (1) участие в биосинтезе, осущ. гидроксилир. при синт. стероидн. горм., желчн. к-т, Ох вит.Д и т.д. (2) участ. в проц. детоксикации: печень и погран. тк. Ген cyp (код. цитохромы): ок. 50 аллелей, опр. субстртн. специф. цитохрома. Полиморфизм цит → подбор дозы лек. ср-в осущ. индивидуально (напр., варфарина). 13 сем-в цитохромов (цифра), п/сем-во (буква), пор. №. Разные субстранты: CYP1A2 – ацетаминофен, CYP2D6 – бета-адреноблокаторы, CYP2E1 – этанол.
(33) Образование активных форм кислорода. Роль АФК в организме, их токсичность. Антиоксидантная система. АФК – это оч реакц/спос мол., сод. делокализованные одиночные неспаренные электроны. Кроме полного четырехэлектронного восстановления молекулы О2 до воды, в дыхательной цепи митохондрий в аэробных клетках всегда происходит и неполное 1-, 2- и 3-электронное восстановление с образованием АФК по реакции:
O2 (+ ē) → •O2¯ (+ ē, 2H+) → H2O2 (+ ē) → HO• (+ ē, 2H+) → 2H2O
Образование АФК происходит либо ферментативным путем, либо неферментативным, когда донорами электрона служат ионы металлов (Fe2+, Сu2+) или убихиноны, а также сами АФК. К числу АФК относят супероксид-радикал(•O2), супероксиданион-радикал (•O2¯), гидроперекисный радикал (НО2•), гидроксил-радикал (НО•), перекись водорода (Н2О2), гипохлорит (ClO¯), синглетный кислород (1О2), NO и пероксинитрит (ONOO¯). АФК генерируются во всех частях клетки. Наибольший вклад вносит дыхательная цепь митохондрий, особенно при низкой концентрации АДФ. Важна роль системы цитохрома Р-450, локализованной в ЭПР. Участвуют ядерная мембрана и другие части клетки, при этом АФК часто возникают не только спонтанно, но и ферментативно (НАДФН-оксидаза дыхательного взрыва в плазматической мембране и ксантиноксидаза в цитоплазме). АФК вызывают образование органических гидропероксидов ДНК, белков, липидов, а также малых молекул. Гидропероксиды химически тоже активны, при последующем метаболизме они переходят в спирты, альдегиды, эпоксиды и другие окисленные соединения. Образование гидропероксидов называют перекисным окислением. Наиболее подвержены действию АФК жирные кислоты, содержащие двойные связи, расположенные через СН2-группу. Именно от этой СН2-группы свободный радикал (инициатор окисления) легко отнимает электрон, превращая липид, содержащий эту кислоту, в свободный радикал. ПОЛ – это цепные реакции, в результате которых образуется большое количество частиц, имеющих неспаренный электрон, которые инициируют дальнейшее распространение перекисного окисления. Состояние, при котором клетка испытывает повышенное воздействие свободных радикалов на фоне недостаточности антиоксидантного потенциала, называется окислительным стрессом. К прооксидантам в живой клетке относятся высокие концентрации кислорода (возникающие, например, при длительной гипербарической оксигенации), ферментные системы, генерирующие супероксидные радикалы (например, ксантиноксидаза, ферменты плазматической мембраны фагоцитов), ионы двухвалентного железа. В состав антиоксидантной системы (далее – АОС) входят как низкомолекулярные АО, так и антиоксидантные ферменты. АОС включает: 1.Ферментативное звено: 1)энзиматические перехватчики, такие как супероксиддисмутаза (СОД), дисмутирующая •О2 до Н2О2, каталаза и глутатионпероксидаза (ГПО), конвертирующие Н2О2 до воды. ГПО вместе с глутатион-S-трансферазой (GSТ) участвует в детоксикации гидропероксидов жирных кислот; 2)ферменты, осуществляющие восст. окисленных низкомол. биоАО (глутатионредуктаза) или участвующие в подд. в функц. активном сост. белковых тиолов (тиоредоксинредуктаза); 3)ферменты, участвующие в подд. внутрикл. стационарного уровня восст. эквивалентов (глюкозо-6-фосфатд ДГ, катализирующая образование НАДФН в пентозофосфатном пути окисления глюкозы); 2.Неферментативное звено: 1)гидрофильные скэвенджеры радикалов: восстановленный глутатион (GSН), аскорбат, урат, тиолы (цистеин, эрготионеин); 2)липофильные перехватчики радикалов: токоферолы, флавоноиды, каротиноиды, убихиноны, билирубин; 3)Антиоксидантные белки, по отдельности относящиеся к ферментативному или неферментативному звену АОС): церулоплазмин, альбумин, ферритин, трансферрин, лактоферрин и др., – участвующие в хранении, транспорте или обезвреживании ионов металлов переменной валентности.
7. ОБМЕН И ФУНКЦИИ УГЛЕВОДОВ |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 142; Нарушение авторского права страницы