
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Общая характеристика организации и функционирования сенсорных систем живых объектов
Сенсорные системы живых объектов, напрамер, человека являются частью его нервной системы, способной воспринимать внешнюю для мозга информацию, передавать ее в мозг и анализировать. Получение информации от окружающей среды и собственного тела является обязательным и необходимым условием существования человека. Термин «сенсорные (лат. sensus — чувство) системы» сменил название «органы чувств», сохранившееся только для обозначения анатомически обособленных периферических отделов некоторых сенсорных систем (как, например, глаз или ухо). Все сенсорные системы состоят из периферических рецепторов, проводящих путей и переключательных ядер, первичных проекционных областей коры и вторичной сенсорной коры. Сенсорные системы организованы иерархически, т. е. включают несколько уровней последовательной переработки информации. Низший уровень такой переработки обеспечивают первичные сенсорные нейроны, которые расположены в специализированных органах чувств или в чувствительных ганглиях и предназначены для проведения возбуждения от периферических рецепторов в центральную нервную систему. Периферические рецепторы — это чувствительные высокоспециализированные образования, способные воспринять, трансформировать и передать энергию внешнего стимула первичным сенсорным нейронам. Центральные отростки первичных сенсорных нейронов оканчиваются в головном или спинном мозге на нейронах второго порядка, тела которых расположены в переключательном ядре. В нем имеются не только возбуждающие, но и тормозные нейроны, участвующие в переработке передаваемой информации. Представляя более высокий иерархический уровень, нейроны переключательного ядра могут регулировать передачу информации путем усиления одних и торможения или подавления других сигналов. Аксоны нейронов второго порядка образуют проводящие пути к следующему переключательному ядру, общее число которых обусловлено специфическими особенностями разных сенсорных систем. Окончательная переработка информации о действующем стимуле происходит в сенсорных областях коры. Существуют сенсорные нейроны, генерирующие потенциалы действия спонтанно, т. е. при отсутствии раздражения (например, сенсорные нейроны вестибулярной системы), такая активность называется фоновой. Частота нервных импульсов в этих нейронах может увеличиваться или уменьшаться в зависимости от интенсивности действующего на вторичные рецепторы стимула, кроме того, она может определяться направлением, в котором отклоняются чувствительные волоски механорецепторов. Например, отклонение волосков вторичных механорецепторов в одну сторону сопровождается повышением фоновой активности сенсорного нейрона, которому они принадлежат, а в противоположную сторону — понижением его фоновой активности. Указанный способ рецепции позволяет получать информацию и об интенсивности стимула, и о направлении, в котором он действует. Рецептивным полем называется область, занимаемая совокупностью всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона. Максимальная величина рецептивного поля первичного сенсорного нейрона определяется пространством, которое занимают все ветви его периферического отростка, а число рецепторов, имеющихся в этом пространстве, указывает на плотность иннервации. Высокая плотность иннервации сочетается, как правило, с малыми размерами рецептивных полей и, соответственно, высоким пространственным разрешением, позволяющим различать стимулы, действующие на соседние рецептивные поля. Малые рецептивные поля типичны, например, для центральной ямки сетчатки и для пальцев рук, где плотность рецепторов значительно выше, чем на периферии сетчатки или в коже спины, для которых характерны большая величина рецептивных полей и меньшее пространственное разрешение. Рецептивные поля соседних сенсорных нейронов могут частично перекрывать друг друга, поэтому информация о действующих на них стимулах передается не по одному, а по нескольким параллельным аксонам, что повышает надежность ее передачи.
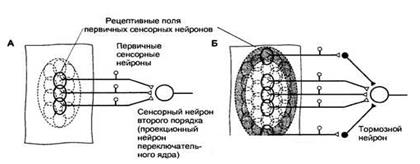
Рисунок 12.2 Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка. А. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов. Б. Раздражение центральной или периферической области рецептивного поля сенсорного нейрона второго и следующих порядков сопровождается противоположным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение с помощью интернейронов переключательного ядра (латеральное торможение). В результате контраста, создаваемого между центром и периферией рецептивного поля, выделяется информация для передачи на следующий иерархический уровень. Величина рецептивных полей сенсорных нейронов второго и следующих порядков больше, чем у первичных сенсорных нейронов, поскольку центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предшествующего уровня. От центра рецептивного поля информация передается непосредственно к сенсорным нейронам следующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, поэтому центр и периферия рецептивного поля являются реципрокными по отношению друг к другу. В результате сигналы от центра рецептивного поля беспрепятственно достигают следующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая функциональная организация рецептивных полей обеспечивает выделение наиболее значимых сигналов, легко различаемых на контрастном с ними фоне. Сенсорные нейроны низшего иерархического уровня способны передавать электрические сигналы одновременно нескольким нейронам следующего уровня благодаря дивергенции к ним своих аксонов. Это повышает надежность передачи информации от одного иерархического уровня к другому и позволяет сохранять чувствительность сенсорной системы при утрате отдельных нейронов. Возбуждение нейронов более высокого иерархического уровня определяется не только действием на них нейронов предыдущего уровня, но и механизмом латерального торможения, существующего в переключательных ядрах (Рисунок 12.3).
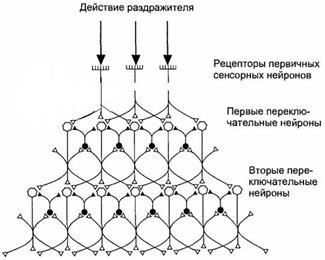
Рисунок 12.3 Схема латерального торможения в сенсорных системах. Степень возбуждения переключательных нейронов определяется интенсивностью раздражения центра или периферии его рецептивного поля. В результате возбуждение будет передаваться от одной части рецептивного поля на фоне торможения или прерывания сигналов от другой его части, что создает эффект контрастирования, способствующий различению действующих стимулов. Большинство раздражителей, воспринимаемых сенсорными системами, оказывает на них комплексное действие, поскольку одновременно возбуждает разные типы рецепторов . Например, прикасающийся к ограниченному участку кожи предмет может быть гладким или шершавым, теплым или холодным, сухим или влажным, а его давление на кожу может усиливаться или ослабевать, он может передвигаться в том или ином направлении. Зрительные ощущения представляют собой сочетание определенной формы воспринимаемого предмета, его движения и комбинации различных цветов, заполняющих ту или иную деталь формы. Сенсорные системы организованы так, что информация о каждом компоненте комплексного раздражителя передается одновременно по нескольким параллельным путям, каждый из которых относится к определенной субмодальности, как, например, форма, движение или цвет наблюдаемого зрительного объекта. В каждом из таких путей независимо друг от друга перерабатываются сигналы, несущие информацию о разных качествах подействовавшего на рецепторы комплексного раздражителя. При этом разные каналы, используемые для передачи сенсорной информации, не дублируют друг друга, в чем проявляется принцип многоканальности, присущий всем сенсорным системам. Высокая упорядоченность нейронных переключений на каждом иерархическом уровне обеспечивает поступление информации от каждого рецептивного поля к соответствующим именно ему кортикальным колонкам проекционной коры. Необходимое для целостного восприятия раздражителя объединение информации, относящейся к разным субмодальностям, происходит в сенсорных регионах коры. Каждая сенсорная система воспринимает действие адекватного стимула в ограниченном диапазоне значений его силы. Наименьший по интенсивности стимул, способный вызвать ощущение, называется абсолютным порогом ощущения. Его величина устанавливается опытным путем, она не одинакова у разных людей и может изменяться у одного и того же человека в зависимости от функционального состояния, различаясь, например, при оптимальной работоспособности и утомлении. Величина абсолютного порога ощущения выше абсолютного порога первичных сенсорных нейронов, так как возбуждение отдельных чувствительных нейронов или их небольшой группы не обязательно приводит к возбуждению высших уровней сенсорной системы вследствие тормозных процессов на предыдущих иерархических уровнях. Поэтому не каждый стимул, возбуждающий рецепторы первичного сенсорного нейрона, субъективно ощущается и осознается. Величина, на которую один стимул надпорогового диапазона должен отличаться от другого, чтобы их разницу можно было субъективно различить, получила название дифференциального порога или порога различения. Ощущаемый прирост интенсивности раздражения (дифференциальный порог) должен превышать ранее действовавший стимул на определенную и постоянную величину. Предложена ступенчатая шкала интенсивности ощущений, в которой за ноль принимается величина абсолютного порога, а зависимость между интенсивностью стимула и ощущением выражается как:
где: Е — интенсивность ощущения, к — постоянная величина, S — интенсивность стимула, Эта зависимость, получившая название закона Вебера—Фехнера, показывает, что линейное увеличение интенсивности ощущения отражает логарифмический рост интенсивности стимула. Закон Вебера—Фехнера характеризует субъективную способность человека различать действующие стимулы (шкала различения), но не оценивает саму интенсивность ощущений, поскольку этот закон основан на допущении равенства любых ее приростов, как к слабым, так и к сильным стимулам. Субъективная оценка интенсивности раздражителя (качественное различие) возрастает при увеличении разницы между пороговой и действующей силой стимула (количественное различие). Однако зависимость между силой стимула и ощущением не одинакова при разной интенсивности стимулов, а потому имеет не линейный, а степенной характер. Для оценки интенсивности ощущений всего диапазона используется шкала Стивенса, устанавливающая зависимость ощущений от силы стимула в виде степенной функции:
где Е — интенсивность ощущения, S — действующая сила стимула, Рецептивные поля различающихся своими рецепторами сенсорных нейронов перекрываются, поэтому при действии на кожу комплекса стимулов одновременно возбуждаются разные виды рецепторов, что позволяет ощущать все динамические и статические свойства такого комплекса. Обработка и анализ информации сигналов от различных рецепторов происходит на высших уровнях сенсорной системы, формирующих комплексное восприятие действующих на поверхность тела стимулов. Плотность механорецепторов в разных участках кожи не одинакова, чем определяются разные показатели пространственного дифференциального порога, т. е. наименьшего расстояния между двумя точками, раздражение каждой из которых ощущается раздельно. |
Последнее изменение этой страницы: 2019-03-22; Просмотров: 147; Нарушение авторского права страницы
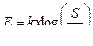 , (12.1)
, (12.1)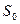 — абсолютный порог.
— абсолютный порог.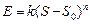 , (12.2)
, (12.2)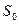 — абсолютный порог, к — константа шкалы, n — показатель степени, который зависит от сенсорной модальности (например, для восприятия светового ощущения он составляет 0,33, громкости звука — 0,6, а для проприоцептивных ощущений — 1,7).
— абсолютный порог, к — константа шкалы, n — показатель степени, который зависит от сенсорной модальности (например, для восприятия светового ощущения он составляет 0,33, громкости звука — 0,6, а для проприоцептивных ощущений — 1,7).