
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ФУНКЦИИ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙСтр 1 из 9Следующая ⇒
СОЕДИНИТЕЛЬНЫЕ ТКАНИ ОБЩИЕ ПРИЗНАКИ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ • развитие в эмбриональном периоде из общего источника – мезенхимы; • высокое содержание межклеточного вещества (которое состоит из волокон и основного аморфного вещества). В некоторых тканях (хрящ, кость) межклеточное вещество играет функционально ведущую роль. ФУНКЦИИ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ • поддержание постоянства внутренней среды организма – гомеостаза, что включает функции: • трофическая (обеспечивает питательными веществами другие ткани); • регуляторная (влияет на деятельность других тканей посредством биологически активных веществ и контактных взаимодействий); • защитная (иммунные реакции и воспаление протекают в соединительной ткани); • опорная, формообразующая - соединительные ткани образуют: - строму различных органов; - капсулы различных органов, связанных со стромальными элементами; - опорные органы (сухожилия, связки, хрящи, кости). • пластическая – участие в восполнении объёма разрушенной части органов и тканей (регенерация).
КЛАССИФИКАЦИЯ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ I. Волокнистые соединительные ткани (собственно соединительные ткани); II. Соединительные ткани со специальными свойствами (жировая, ретикулярная, пигментная, слизистая); III .Скелетные соединительные ткани (хрящевые и костные).
ВОЛОКНИСТЫЕ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ характеризуются высоким содержанием межклеточного вещества (матрикса), состоящего из двух компонентов: • волокон, которые выполняют функциональную роль; • основного аморфного вещества, которое заполняет пространства между волокнами и клетками. Классификация волокнистых соединительных тканей основана на соотношении клеток и межклеточного вещества, а также степени упорядоченности волокнистого компонента. Рыхлая волокнистая соединительная ткань (РВСТ). Для этого типа ткани характерно: • сравнительно невысокое содержание волокон в межклеточном веществе (1), • относительно большой объем основного аморфного вещества (2) • многочисленный и разнообразный клеточный состав (3). Плотная волокнистая соединительная ткань. Для этого типа ткани характерно: • преобладание в межклеточном веществе волокон; • незначительный объем основного аморфного вещества; • малочисленный и однообразный клеточный состав. Различают два подтипа плотной волокнистой соединительной ткани: • оформленная плотная волокнистая соединительная ткань (все волокна ориентированы в одном направлении и образуют параллельные пучки, как в сухожилиях, или переплетаются в одной плоскости, как в апоневрозах); • б) неоформленная плотная волокнистая соединительная ткань (волокна ориентированы случайным образом, образуя трёхмерную сеть переплетающихся волокон, как в сетчатом слое кожи или капсулах органов). Фиброцит • конечная форма дифферона; • малоактивная клетка; • долгоживущая клетка; • неспособна к пролиферации; • узкая, веретенообразная форма, с тонкими отростками; • ядро плотное; • синтетический аппарат развит слабо; • много лизосом; Функция фиброцитов: регуляция метаболизма и поддержание стабильности межклеточного вещества. Фиброкласты - клетки, которые специализируются на разрушении межклеточного вещества; • обеспечивают перестройку межклеточного вещества; • многочисленны в молодой соединительной ткани (грануляционной) и рубцах; • для цитоплазмы фиброкластов характерны вакуоли с коллагеновыми фибриллами на разных стадиях лизиса; • расщепление коллагена происходит вне- и внутриклеточно. Миофибробласты – особый тип фибробластов: • более половины их цитоплазмы занимают элементы сократительного аппарата (актин); • активно участвуют в репаративных процессах • контракция раны: сокращаясь, они стягивают края раны и образуют коллаген, который заполняет поврежденный участок.
МАКРОФАГИ • Вторые по численности клетки РВСТ; • Происхождение - потомки СКК, образуются из моноцитов крови; • Особенно многочисленны в собственной пластинке слизистых и серозных оболочек. • Активные макрофаги обладают высокой подвижностью, изменчивой, обычно отростчатой формой (микровыросты, псевдоподии) с неровными, но чёткими краями. • Ядра темнее, чем у фибробластов. • Для цитоплазмы макрофагов характерно: многочисленные лизосомы и крупные фаголизосомы, пиноцитозные пузырьки, развитые элементы цитоскелета; остальные органеллы развиты умеренно. Функции макрофагов • Фагоцитоз - распознавание, захват и переваривание поврежденных, зараженных, опухолевых и погибших клеток, компонентов межклеточного вещества, экзогенных микроорганизмов и субстанций (на поверхности имеются рецепторы для иммуноглобулинов, антигенов опухолевых клеток). Различают фагоцитоз неспецифический и специфический. • неспецифический фагоцитоз характерен для лёгочных макрофагов, захватывающих частицы пыли, сажи и т д. • специфический фагоцитоз – сначала иммуноглобулины и белки комплемента плазмы крови (объединенные названием опсонины) окружают (опсонизируют) бактерию. Макрофаг имеет рецепторы к опсонинам и легко захватывает опсонизированные бактерии и образует фагосомы. Лизосомы содержат лизоцим, разрушающий бактериальную стенку, и гидролитические ферменты. • Макрофаги могут секретировать содержимое лизосом в инфицированных зонах. • Индукция иммунных реакций – играют роль антиген-представляющих клеток; макрофаги осуществляют процессинг антигенного материала - эпитопы антигенов вместе с молекулами главного комплекса гистосовместимости выделяются на поверхность клетки – узнавание иммунокомпетентными клетками. • Регуляция деятельности других типов (фибробластов, лимфоцитов, тучных клеток и др.). Макрофаги секретируют ряд биоактивных факторов, которые называются монокины: такие как интерлейкин-1; фактор хемотаксиса нейтрофилов; эндогенные пирогены (через центр терморегуляции вызывают повышение температуры); интерферон (противовирусное); антибактериальные: лизоцим, активные метаболиты кислорода. • Участие в противоопухолевом иммунитете. Макрофаги вырабатывают фактор некроза опухолей, который оказывает цитотоксическое действие на трансформированные клетки. • Регуляция тканевого гомеостаза: макрофаги элиминируют старые элементы тканей, участвуют в тканевом обмене веществ (особенно обмене жиров), регулируют состояние межклеточного вещества • Регуляция регенерации: секретируют ряд веществ, стимулирующих заживление ран; Участвуют в макрофагической фазе воспаления (фагоцитируют разрушенные части тканей, бактерии и выделяют факторы, стимулирующие фибробласты). В очаге повреждения макрофаги могут превращаться в особые виды – гигантские многоядерные клетки и эпителиоидные клетки. Функции тучных клеток Регуляторная: поддержание гомеостаза соединительной ткани (путём медленного выделения малых доз биоактивных веществ, которые влияют на проницаемость и тонус сосудов и поддержание баланса жидкости в тканях); Защитная: при воспалении происходит быстрое, локальное выделение медиаторов воспаления и хемотаксических факторов, привлекающих нейтрофилы и эозинофилы
Участие в аллергических реакциях. Макрофаги имеют рецепторы к иммуноглобулинам класса Е (IgE – образуются в ответ на проникновение антигенов-аллергенов) на плазмолемме → выделение содержимого гранул (гистамин) и синтез ряда новых веществ (простагландины, тромбоксан и т.д). При анафилактической дегрануляции гранулы сливаются в цепочки, образуя внутрицитоплазматический канал, происходит массированное выделение СГ → быстрое сосудорасширяющее действие на капилляры и венулы, повышает их проницаемость и выход плазмы в ткани, спазм гладких мышц бронхиол, повреждение эпителия. (бронхоспазм, острый ринит, отеки, зуд, понос, падение кровяного давления – анафилактический шок). Вещества, угнетающие дегрануляцию тучных клеток, с различными механизмами фармакологического действия широко распространены в качестве профилактики и лечения аллергических реакций. ЖИРОВЫЕ КЛЕТКИ (адипоциты) образуются из юных фибробластов путем накопления в цитоплазме мелких липидных капель, которые сливаются в одну крупную (однокапельные адипоциты).
Локализация: встречаются повсеместно, в виде скоплений (дольки) или по отдельности, вдоль сосудов. Морфология: Крупные клетки сферической формы, с уплощенным ядром, тонким ободком цитоплазмы с органеллами. Функция: Адипоциты – клетки с высокой метаболической активностью: • обмен липидов, углеводов; • депо жирорастворимых витаминов, • вырабатывают ряд гормонов (например, лептин – регулятор потребления пищи - и эстрогены) • депо стероидных гормонов.
ЛЕЙКОЦИТЫ. Из крови в РВСТ попадают все виды лейкоцитов: гранулоциты, лимфоциты, моноциты, которые превращаются здесь в макрофаги. Эти «пришлые» клетки осуществляют свои защитные реакции (воспаление, иммунный ответ) в соединительной ткани. ПЛАЗМОЦИТЫ образуются из В-лимфоцитов; Клетки округлой или овальной формы; Имеют эксцентрично расположенное ядро с характерным расположением хроматина (вид «колеса со спицами»), около которого находится комплекс Гольджи и центриоли – «дворик». Базофильная цитоплазма содержит хорошо развитую гранулярную эндоплазматическую сеть, в которой синтезируются белки-антитела. Функции плазматических клеток . Плазмоциты - эффекторные клетки гуморального иммунитета, единственные клетки в которых происходит синтез иммуноглобулинов – антител. ПИГМЕНТНЫЕ КЛЕТКИ (меланоциты) имеют нейральное происхождение (из нервного гребня). Морфология. Клетки имеют отростчатую форму и содержат пигменты меланины. Функция. Важнейший защитный эффект меланина – способность поглощать свободные радикалы. Пигментоциты принимают также участие в регуляции тонуса кровеносных сосудов (выделяют биоактивные амины). Пигментные клетки численно преобладают и играют ведущую функциональную роль в специальном типе соединительной ткани - пигментной ткани (радужка и сосудистая оболочка глаза). o АДВЕНТИЦИАЛЬНЫЕ КЛЕТКИ Адвентициальные клетки – малодифференцированные клетки, имеют уплощенную форму, слабобазофильную цитоплазму, овальное ядро. Локализуются в наружной оболочке мелких кровеносных сосудов. Дифференцируясь, Адвентициальные клетки способны превращаться в фибробластоциты, миофибробласты и жировые клетки. o ПЕРИЦИТЫ - клетки отростчатой формы в стенке гемокапилляра. Участвуют в регуляции просвета капилляров и в трофике. МЕЖКЛЕТОЧНОЕ ВЕЩЕСТВО РВСТ КОЛЛАГЕНОВЫЕ ВОЛОКНА Коллагеновые волокна образованы белками коллагенами, которые являются наиболее распространенными белками организма. Характерная структура молекулы коллагена - длинная жесткая тройная спираль, которая состоит из 3-х скрученных спирально полипептидных цепей (α-цепей). Известны более 30 вариантов цепей, различных по химическому составу (типичные аминокислоты α-цепи: глицин, пролин, лизин, гидроксипролин). Варианты дают около 19 типов коллагена. Наиболее значение имеют пять первых типов (I - V ). Коллагены I, II, III и Y типов являются фибриллярными, то есть они способны формировать нитевидные структуры - филаменты и фибриллы. Остальные коллагены, включая IY тип, этой способностью не обладают и являются аморфными (образуют плоские сети). Помимо фибробластов, коллаген могут синтезировать остеобласты, хондробласты, одонтобласты, цементобласты, ретикулярные клетки, гладкие миоциты. Коллаген I типа характерен для рыхлой волокнистой соединительной ткани, дермы, костей, дентина, цемента, связок, сухожилий; Коллаген II входит в состав хрящи, стекловидное тело глаза; Коллаген III – ретикулярные волокна; Коллаген IY – (нефибриллярный) образует базальные мембраны, капсулу хрусталика; Коллаген Y – определяется в мышечных базальных мембранах, в стенках кровеносных сосудов, в коже, дентине. Биосинтез коллагена включает внутриклеточный и внеклеточный этапы. · Внутриклеточный этап включает: поглощение и транспорт необходимых аминокислот → синтез полипетидных α-цепей, посттрансляционные изменения, сборка трех α-цепей и образование проколлагена (в просвете грЭПС) → перенос в комплекс Гольджи, где происходит терминальное гликозилирование и упаковка в секреторные пузырьки → экзоцитоз молекул проколлагена в межклеточную среду. · Внеклеточный этап включает отщепление регистрационных пептидов с помощью специфических протеаз, связанных с плазмолеммой и образование молекул тропоколлагена толщиной 1,5 нм – нерастворимых, способных к самосборке. Далее происходит упорядоченная внеклеточная агрегация коллагеновых фибрилл толщиной 20-120 нм. Молекулы тропоколлагена связываются в продольные цепочки (микрофибриллы), располагаясь параллельно друг другу. При этом внутри каждой цепочки есть зоны зазора (промежутки); при этом в соседних цепочках молекулы тропоколлагена сдвинуты друг относительно друга на четверть длины. При окраске выявляется поперечная исчерченность (период 64-68 нм) коллагеновых фибрилл вследствие отложения красителя в зонах зазора. Коллагеновые фибриллы образуют коллагеновые волокна толщиной 1-20 мкм, которые могут объединяться в коллагеновые пучки (первичные, вторичные, третичные) Морфология . Коллагеновые волокна - оксифильные, продольно исчерченные, извитые тяжи, лежат в ткани поодиночке или образуют пучки. Основные функции коллагеновых волокон: обеспечивают высокие механические свойства ткани (поскольку эти волокна - прочные и нерастяжимые); обеспечивают структурную организацию (архитектонику) соединительной ткани; обеспечивают взаимодействие между клетки и межклеточным веществом; влияют на пролиферацию, дифференцировку, миграцию и функциональную активность различных клеток. Поскольку ферментное гликозилирование пролина и лизина зависит от витамина С, наряду с другими симптомами, авитаминоз (цинга) характеризуется расшатыванием и выпадением зубов, из-за нарушения обновления коллагеновых волокон периодонтальной связки – главного элемента поддерживающего аппарата зуба. РЕТИКУЛЯРНЫЕ ВОЛОКНА Основа ретикулярных волокон – коллаген III типа. Ретикулярные волокна имеют малый диаметр (0.1-2 мкм) и образуют тонкие трехмерные сети. Каждое волокно образовано пучком коллагеновых микрофибрилл, заключенных в оболочку из гликопротеинов и протеогликанов. Из-за такой оболочки ретикулярные волокна, в отличие от коллагеновых, не окрашиваются эозином, а окрашиваются солями серебра, поэтому их часто называют аргирофильными волокнами. Функция ретикулярных волокон - опорная: они образуют каркас миелоидной и лимфоидной тканей, оплетают базальные мембраны, окружают капилляры и нервные волокна, гладкомышечные клетки, образуют поддерживающую строму гепатоцитов. ЭЛАСТИЧЕСКИЕ ВОЛОКНА Эластические волокна наиболее многочисленны в участках, обладающих подвижностью: в подслизистой основе пищеварительного канала, в стенках артерий. Толщина эластических волокон 0.2-10 мкм; эти волокна ветвятся и анастомозируют друг с другом, образуя сети. В рыхлой соединительной ткани эластические волокна не образуют пучков. Функция эластических волокон – обеспечение способности к обратимой деформации. Главный белковый компонент эластических волокон – белок эластин. Молекулы эластина имеют в состоянии покоя форму скрученных нитей. При растяжении молекулы распрямляются, после нагрузки – закручиваются. Молекулы ковалентно сшиты в комплексы, формируют волокна и пластины (мембраны – в артериях). Структура на электронно-микроскопическом уровне: в зрелом эластическом волокне выявляется центральный светлый аморфный компонент (эластин) и периферический микрофибриллярный компонент (тонкие волоконца гликопротеина фибриллина). Кроме зрелых собственно эластических волокон в эластическую систему входят волокна с меньшей степенью зрелости: окситалановые, образованные микрофибриллами, сходными с периферическим компонентом зрелого волокна; элауниновые, более зрелые, которые имеют островки микрофибрилл среди аморфного вещества. По мере зрелости (в ходе эластогенеза) на фибриллярный компонент откладывается эластин, эластин (Э) накапливается, а микрофибриллярный компонент (МФК) постепенно оттесняется на периферию и разрушается. ОСНОВНОЕ АМОРФНОЕ ВЕЩЕСТВО Основное аморфное вещество заполняет промежутки между волокнами и окружает клетки. Имеет аморфное строение, прозрачно, базофильно, с низкой электронной плотностью. На молекулярном уровне состоит из макромолекулярных гидратированных комплексов протеогликанов и структурных гликопротеинов. Протеогликаны состоят из пептидной цепи (так называемый сердцевинный, или осевой белок), связанной с гликозаминогликанами. Гликозамингликаны (ГАГ) – крупные (до 200 сахаров в цепи),
Основные особенности ГАГ заключаются в том, что: · ГАГ очень гидрофильны и, соединяясь с водой, могут образовывать подвижное гелеподобнное вещество, упругое как резина; · Даже при низкой концентрации ГАГ образуются гели, занимающие большой объём. Эта способность к набуханию позволяет матриксу противостоять сжимающим силам. Например, когда вы подпрыгиваете, ГАГ коленных и голеностопных суставов амортизируют силу удара, получаемого в момент приземления. · Коллагеновые волокна, погруженные в гелевый матрикс, обеспечивают его прочность подобно стальным стержням, укрепляющим бетон. · Благодаря высокой плотности отрицательного заряда они прочно связывают катионы. · ГАГ формируют буферную среду. Функциональная роль протеогликанов: · играют важную роль в транспорте электролитов и воды; · связывают и накапливают факторы роста; · взаимодействуют с молекулами коллагена и способствуют их правильной укладке; · обеспечивают связь между поверхностью клеток и компонентами межклеточного вещества. Структурные гликопротеины: фибронектин, ламинин, энтактин и другие - в отличие от протеогликанов в своей основе имеют разветвленную пептидную цепь, с которой связано небольшое количество простых гексоз. Роль структурных гликопротеинов: · организация межклеточного вещества; · посредники во взаимодействиях между клетками и компонентами межклеточного вещества; · образование базальных мембран. Фибронектин обеспечивает связь клеток с внеклеточным матриксом (коллагеном и ГАГ) через трансмембранные белки плазмолеммы клеток (цепочка: коллаген/ГАГ – гепаран сульфат – фибронектин - интегрины – актиновые филаменты цитоскелета); обеспечивает прикрепление к нему фибробластов и других клеток, влияя на их функции, подвижность. Ламинин – важнейший компонент базальной мембраны, связывает через молекулы клеточной адгезии клеточной мембраны базальную плазмолемму и IY коллаген («сшивает» клетку с базальной мембраной). Энтактин, другой нефибриллярный гликопротеин, связывает («сшивает») ламинин с коллагеном 4-го типа. ВОСПАЛЕНИЕ Воспаление - защитно-приспособительная реакция на местное повреждение, выработанная в ходе эволюции. Факторы, вызывающие воспаление, могут быть экзогенными (инфекция, травма, ожог, гипоксия) или эндогенными (очаг некроза, отложение солей). Биологический смысл этой защитной реакции - ликвидация или ограничение от здоровой ткани очага повреждения, и регенерация ткани. Хотя это и защитная реакция, но в некоторых случаях проявления этой реакции, особенно хронического воспаления, способны вызвать тяжелые повреждения тканей. Фазы воспаления: I . фаза альтерации – повреждение тканей и выделение медиаторов воспаления, комплекса биоактивных веществ, отвечающих за возникновение и поддержание воспалительных явлений. Медиаторы воспаления: гуморальные (из плазмы крови) – кинины, факторы свертывания и т.д.; клеточные медиаторы выделяются клетками в ответ на повреждение; вырабатываются моноцитами, макрофагами, тучными клетками, гранулоцитами, лимфоцитами, тромбоцитами. Эти медиаторы: биоамины (гистамин, серотонин), эйкозаноиды (производные арахидоновой кислоты: простагландины, лейкотриены), и другие. II . фаза экссудации включает: · изменения микроциркуляторного русла: спазм артериол, затем расширение артериол, капилляр и венул – возникает гиперемия – покраснение и повышение температуры. · формирование жидкого (бесклеточного) экссудата – благодаря повышению проницаемости сосудов, изменения осмотического давления в очаге воспаления (из-за повреждения) и гидростатического в сосудах. Нарушение оттока приводит к возникновению отёка. · формирование клеточного экссудата (миграция лейкоцитов через эндотелий). Клеточный состав фаз воспаления: 1 фаза: на начальных этапах наиболее активно выселяются нейтрофильные гранулоциты, которые выполняют фагоцитарную и микробицидную функции; в результате их активности образуются продукты распада, которые привлекают в очаг воспаления моноциты, выселяющие из крови; 2 фаза: моноциты в соединительной ткани превращаются в макрофаги. Макрофаги фагоцитируют погибшие нейтрофилы, клеточный детрит, микроорганизмы и могут инициировать иммунный ответ. В очаге хронического воспаления преобладают микрофаги и лимфоциты, которые образуют скопления – гранулёмы. Сливаясь, макрофаги образуют гигантские многоядерные клетки. III . фаза пролиферации (репарации) – Макрофаги, лимфоциты и другие клетки вызывают: хемотаксис, пролиферацию и стимуляцию синтетической активности фибробластов; активацию образования и роста сосудов. Образуется молодая грануляционная ткань, откладывается коллаген, формируется рубец.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ ЖИРОВАЯ ТКАНЬ Жировая ткань представляет собой особую разновидность соединительной ткани, в которой основной объём занимают жировые клетки – адипоциты. Жировая ткань повсеместно распространена в организме, составляя 15-20% массы тела у мужчин и 20-25% - у женщин (т.е. 10-20 кг у здорового человека). При ожирении (а в развитых странах это около 50% взрослого населения) масса жировой ткани увеличивается до 40-100 кг. Аномалии содержания и распределения жировой ткани связаны с рядом генетических нарушений и эндокринных расстройств. У млекопитающих, включая человека, имеются два вида жировой ткани – белая и бурая, которые различаются по цвету, распределению в организме, метаболической активности, строению образующих их клеток (адипоцитов) и степени кровоснабжения. Белая жировая ткань – преобладающий вид жировой ткани. Образует поверхностные (гиподерма – слой подкожной жировой клетчатки) и глубокие – висцеральные – скопления, образует мягкие упругие прослойки между внутренними органами. В эмбриогенезе жировая ткань развивается из мезенхимы. Предшественники адипоцитов – малодифференцированные фибробласты (липобласты), лежащие по ходу мелких кровеносных сосудов. В ходе дифференцировки в цитоплазме образуются сначала мелкие липидные капли, капли сливаются друг с другом, образуя одну крупную каплю (95-98% объёма клетки), а цитоплазма и ядро смещаются к периферии. Такие жировые клетки называются однокапельными адипоцитами. Клетки утрачивают отростки, приобретают сферическую форму, в ходе развития их размер увеличивается в 7-10 раз (до 120 мкм в диаметре). Цитоплазма характеризуется развитой агранулярной ЭПС, мелким комплексом Гольджи, небольшим количеством митохондрий.
Белая жировая ткань состоит из долек (компактных скоплений адипоцитов), разделенных тонкими прослойками рыхлой волокнистой соединительной ткани, несущими кровеносные и лимфатические сосуды и нервы. В дольках клетки приобретают форму многогранников. Функции белой жировой ткани: · энергетическая (трофическая): адипоциты обладают высокой метаболической активностью: липогенез (отложение жиров) - липолиз (мобилизация жиров) – обеспечение организма резервными источниками; · опорная, защитная, пластическая – полностью или частично окружает различные органы (почки, глазное яблоко и.т.д.). Резкое похудание может привести к смещению почек; · теплоизолирующая; · регуляторная – в процессе миелоидного кроветворения адипоциты входят в состав стромального компонента красного мозга, создающего микроокружение для пролиферирующих и дифференцирующихся клеток крови; · депонирующая (витамины, стероидные гормоны, вода) · эндокринная – синтезирует эстрогены (главный источник у мужчин и пожилых женщин) и гормон, регулирующий потребление пищи – лептин. Лептин тормозит секрецию гипоталамусом особого нейропептида NPY, который усиливает потребление пищи. При голодании секреция лептина снижается, при насыщении – возрастает. Недостаточная выработка лептина (или отсутствие рецепторов к лептину в гипоталамусе) ведёт к ожирению. Ожирение В 80% увеличение массы жировой ткани происходит вследствие нарастания объёма (гипертрофии) адипоцитов. В 20% (при наиболее тяжелых формах ожирения, развивающихся в молодом возрасте) – увеличение числа адипоцитов (гиперплазия): число адипоцитов может увеличиться в 3-4 раза. Голодание Снижение массы тела в результате лечебного или вынужденного голодания сопровождается падением массы жировой ткани – усиление липолиза и угнетение липогенеза – резкое уменьшение объемов адипоцитов при сохранении их общего числа. При возобновлении нормального питания клетки быстро накапливают липиды, клетки увеличиваются в размерах, и превращаются в типичные адипоциты, в результате чего происходит быстрое восстановление массы тела после отмены диеты. Жировая ткань на ладонях, подошвах и в ретроорбитальных участках очень устойчива к процессам липолиза. Снижение массы жировой ткани более чем на треть от нормы, вызывает дисфункцию системы гипоталамус-гипофиз-яичники – подавление менструального цикла и бесплодие. Нервная анорексия - один из видов пищевых расстройств, при котором запас жировой ткани снижается до 3 % нормального уровня массы жировой ткани, нередко заканчивается смертельным исходом. Бурая жировая ткань У взрослого человека бурая жировая ткань присутствует в небольшом количестве, лишь в нескольких, чётко очерченных участках (между лопаток, на задней поверхности шеи, в воротах почек). У новорожденных она составляет до 5% массы тела. Её содержание мало меняется при недостаточном или избыточном питании. Бурая жировая ткань наиболее сильно развита у животных, впадающих в зимнюю спячку. Особенности строения
Адипоциты бурой ткани более мелкие в сравнении с адипоцитами белой жировой ткани клетки, полигональной формы. Ядро располагается в центре клетки, характерны множественные жировые капли различных размеров, поэтому клетки бурой жировой ткани называются многокапельными адипоцитами. Значительный объем цитоплазмы занимают многочисленные митохондрии с развитыми ламеллярными кристами. Дольки бурой жировой ткани разделены очень тонкими прослойками рыхлой волокнистой соединительной ткани, но кровоснабжение очень обильное. Терминали симпатических нервных волокон погружены в участки цитоплазмы адипоцитов. Буровато-красный цвет этого типа жировой ткани связан с густой сетью капилляров в ткани, а также высоким содержанием окрашенных окислительных ферментов – цитохромов – в митохондриях адипоцитов. Ведущая функция бурой жировой ткани – термогенез , продукция тепла. На кристах митохондрий адипоцитов этой ткани мало оксисом (участок расположения АТФ-синтетического комплекса). Митохондрии содержат особый белок – UCP (uncoupling protein – разобщающий белок), или термогенин, благодаря которому в результате окисления жиров энергия не запасается в виде макроэргических соединений (АТФ), а рассеивается в виде тепла. Окислительная способность многокапельных адипоцитов в 20 раз выше, чем у однокапельных адипоцитов. Обильное кровоснабжение обеспечивает быстрое отведение вырабатываемого тепла. С током крови тепло распространяется по всему телу. Главным фактором, вызывающим термогенез и мобилизацию липидов из бурой ткани, является стимуляция симпатической нервной системы, адреналин, норадреналин. Ретикулярная ткань
Ретикулярная ткань - специализированная соединительная ткань, которая входит в качестве структурной основы (стромы) в состав кроветворных тканей – миелоидной и лимфоидной. Её элементы – ретикулярные клетки и ретикулярные волокна образуют трехмерную сеть, в петлях которой развиваются клетки крови. Ретикулярные клетки – крупные, отростчатые, фибробластоподобные клетки, формирующие сеть. Для них характерно округлое светлое ядро с крупным ядрышком, слабооксифильная цитоплазма. Отростки ретикулярных клеток связаны между собой щелевыми контактами. Функции ретикулярной ткани: · поддерживающая; · создание микроокружения в миелоидной ткани: транспорт питательных веществ; секреция гемопоэтинов – гуморальных факторов, регулирующих деление и дифференцировку клеток крови; адгезивные контакты с развивающимися клетками крови. · синтетическая: образуют ретикулярные волокна и основное аморфное вещество. · барьерная: контроль миграции форменных элементов в просвет сосудов. Ретикулярные волокна образованы коллагеном III типа, оплетают ретикулярные клетки, в некоторых участках оказываются охваченными цитоплазмой этих клеток. Волокна довольно тонкие (до 2 мкм), обладают аргирофилией (окрашиваются солями серебра) и дают ШИК-PAS реакцию (реактив Шиффа–йодная кислота, выявляет соединения, богатые углеводными группами), поскольку ретикулярные микрофибриллы покрыты оболочкой из гликопротеинов и протеогликанов. Основное вещество – протеогликаны и гликопротеины связывают, накапливают и выделяют факторы роста, влияющие на процессы гемопоэза. Структурные гликопротеины ламинин, фибронектин и гемонектин способствуют адгезии кроветворных клеток к строме. Кроме ретикулярных клеток, в ретикулярной ткани присутствуют макрофаги и дендритные антиген-представляющие клетки. Пигментная ткань Пигментная ткань близка по строению к рыхлой волокнистой соединительной ткани, однако содержит значительно большее количество пигментных клеток. Пигментная ткань образует радужку и сосудистую оболочку глаза. Пигментные клетки подразделяются на меланоциты и меланофоры. Меланоциты – отростчатые клетки, контактирующие с другими клетками этой ткани. Цитоплазма содержит развитый синтетический аппарат и большое количество меланосом – гранул, содержащих тёмный пигмент меланин. Эти клетки синтезируют меланин. Меланофоры – имеют слабо развитый синтетический аппарат и значительное число зрелых меланиновых гранул. Эти клетки не синтезируют, а только поглощают готовые меланиновые гранулы. Другие клетки, встречающиеся в пигментной ткани: фибробласты, фиброциты, макрофаги, тучные клетки, лейкоциты. Функции пигментной ткани: защита от повреждающего и мутагенного действия ультрафиолета, поглощение избытка световых лучей. Слизистая ткань Видоизмененная рыхлая волокнистая соединительная ткань с резким преобладанием межклеточного вещества, в котором волокнистый компонент развит слабо. Слизистая ткань имеет гелеподобную консистенцию. В ней отсутствуют сосуды и нервные волокна. Слизистая ткань заполняет пупочный канатик плода (так называемый Вартонов студень). Близкое строение имеет стекловидное тело глазного яблока. Клетки слизистой ткани сходны с фибробластами, но содержат много гликогена в цитоплазме. В межклеточном веществе резко преобладает однородное и прозрачное основное вещество. Высокое содержание гиалуроновой кислоты в основном веществе, создает значительный тургор, что препятствует сдавливанию пупочного канатика. Хрящевые ткани Хрящевые ткани развиваются из склеротомной мезенхимы. Хрящевые ткани входят в состав органов дыхательной системы (нос, гортань, трахея, бронхи), ушной раковины, суставов, межпозвоночных дисков. Общие свойства хрящевых тканей: · сравнительно низкий уровень метаболизма; · отсутствие сосудов; · прочность и эластичность; клетки хрящевой ткани лежат в лакунах, окруженных плотным межклеточным веществом. Классификация хрящевых тканей основана на особенностях строения и биохимического состава их межклеточного вещества (матрикса). На основе этих различий выделяют три вида хрящевых тканей: · гиалиновая хрящевая ткань · эластическая хрящевая ткань · волокнистая хрящевая ткань. Гиалиновая хрящевая ткань Наиболее распространенный вид хрящевых тканей - гиалиновая хрящевая ткань. Этот вид получил название вследствие того, что на макропрепарате имеет внешнее сходство с матовым стеклом (греч.hyalos – стекло). Клетки хрящевой ткани - хондроциты – высокоспециализированные клетки, вырабатывающие межклеточное вещество ткани. Располагаются в лакунах поодиночке или в виде изогенных групп (до 8-12 клеток). Изогенные группы – потомки одной клетки, они образуются в результате деления молодых хондроцитов, но не могут покинуть лакуну, так как матрикс уже достаточно плотный. Для хондроцитов характерны светлое ядро с ядрышком, в цитоплазме имеется гранулярная эндоплазматическая сеть, комплекс Гольджи, включения гликогена и липидные. Хондроциты синтезируют коллаген II типа, сульфатированные гликозаминогликаны (хондроитин-сульфат и др.) и гликопротеины. Матрикс гиалинового хряща представлен: · коллагеном 2-го типа, который образует тонко-волокнистый каркас; · протеогликанами, которые формируют агрегаты и взаимодействуют с коллагеновыми волокнами; · интерстициальной водой (до 65-85% его веса), которая может перемещаться по матриксу, и благодаря своей низкой сжимаемости, обеспечивает упругость ткани. Коллаген 2-го типа образует тонкие фибриллы, собирающиеся в волокна, расположенные по направлению сил, воздействующих на хрящ. На препаратах матрикс кажется однородным, гомогенным, поскольку коллагеновые волоконца очень тонкие, на пределе разрешения светового микроскопа, и не различимы на фоне основного вещества матрикса, имеющего сходный коэффициент преломления. Кроме коллагена 2-го типа, в составе межклеточного вещества гиалинового хряща содержатся коллагены IX типа (сшивает коллагеновые волокна 2-го типа), X типа (с ним связана способность хряща к обызвествлению – отложению неорганических солей).
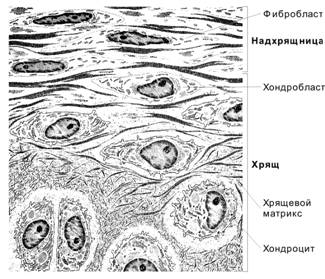
Аморфное вещество хрящевых тканей представлено в основном протеогликанами, образующими агрегаты. В состав протеогликанов входят гликозаминогликаны (80-90%), которые под прямым углом присоединяются к осевому белку, образующему стержень протеогликана. В результате формируется структура, напоминающая ёршик для мытья пробирок или ламповую щётку. Такие мономеры протеогликанов (до 80 субъединиц) через связующие белки соединяются с очень длинной линейной молекулой гиалуроновой кислоты, образуя агрегаты и суперагрегаты протеогликанов. Протеогликаны способны связывать огромные количества воды. Адгезивные гликопротеины (хондронектин) связывают агрегаты и коллагеновые волокна друг с другом и поверхностью хондроцитов в целостную систему, обеспечивая прочность хрящевой ткани. Межклеточное вещество (матрикс) хряща подразделяют на две зоны: территориальный матрикс и интертерриториальный матрикс. Территориальный матрикс непосредственно окружает изогенные группы клеток, и состоит из капсулы, окружающей лакуну (коллаген IX типа) и протеогликанов. Территориальный матрикс окрашивается базофильно. Интертерриториальный матрикс находится между лакунами, представляет собой более старые участки межклеточного вещества, здесь преобладают коллагеновые волокна, и этот матрикс окрашивается слабо базофильно или даже оксифильно. В целом, матрикс хряща обеспечивает прочность и упругость ткани. Гиалиновый хрящ рёбер, гортани, воздухоносных путей окружен надхрящницей, из сосудов которой он получает питание. Надхрящница состоит из двух слоёв: · наружного фиброзного, образованного плотной волокнистой соединительной тканью; · внутреннего хондрогенного, образованного рыхлой волокнистой соединительной тканью, содержащей хондрогенные клетки.
Функции надхрящницы: · трофическая – в ней находятся кровеносные сосуды, которые обеспечивают питание хряща; · регенераторная - внутренний хондрогенный слой содержит прехондробласты, которые при стимуляции могут превращаться в хондробласты, активно синтезирующие матрикс и обеспечивающие рост и восстановление хряща; · опорно-механическая, обеспечивает связь хряща с другими анатомическими структурами (сухожилиями, связками и др.). Под надхрящницей расположена зона малодифференцированного хряща, в которой молодые хондроциты лежат в лакунах поодиночке параллельно надхрящнице. Суставный гиалиновый хрящ благодаря гладкой поверхности обеспечивает скольжение костей друг относительно друга, а его упругие свойства амортизируют всевозможные удары. Суставный хрящ не имеет надхрящницы, и его питание идёт в основном из синовиальной жидкости. Здоровый хрящ выделяет так называемый антиангиогенный фактор, препятствующий врастанию в хрящевую ткань кровеносных сосудов. При старении хряща интенсивность выработки этого фактора снижается. Врастание кровеносных сосудов способствует минерализации хряща и превращению его в кость. За счёт пре- и хондробластов надхрящницы происходит рост хряща с периферии - аппозиционный рост. Кроме того, в молодом хряще хондроциты способы к делению и синтезу межклеточного вещества. За счёт их деления и отложения нового матрикса происходит рост хряща изнутри - интерстициальный рост.
Эластическая хрящевая ткань Эластическая хрящевая ткань входит в состав хрящей ушной раковины, надгортанника, стенок бронхов среднего калибра. Эта хрящевая ткань обеспечивает гибкость и эластичность - обратимую деформацию структур, в состав которых он входит. По своему строению эластический хрящ похож на гиалиновый хрящ рёбер и воздухоносных путей. Основное отличие эластической хрящевой ткани от гиалинового хряща состоит в том, что в составе её межклеточного вещества преобладают тонкие эластические волокна (90%), которые идут в разных направлениях и образуют густую сеть. Поскольку в составе межклеточного вещества эластического хряща отсутствует коллаген Х типа, эластический хрящ никогда не минерализуется. Содержание основного вещества незначительно. Волокнистая хрящевая ткань Волокнистая хрящевая ткань входит в состав межпозвоночных дисков, лонного сращения, встречается в местах переходов сухожилий и связок в гиалиновый хрящ. Эта ткань никогда не встречается изолированно, а служит «мостиком», связывающим плотную волокнистую соединительную ткань и гиалиновый хрящ. Хондроциты волокнистого хряща лежат в лакунах поодиночке или в виде мелких изогенных групп и выстраиваются в цепочки вдоль толстых пучков волокон коллагена I типа (90%), который синтезируют клетки этого хряща в дополнении к коллагену II типа. Вследствие такого состава матрикс волокнистого хряща окрашивается эозинофильно. При переходе к сухожилию хондроциты постепенно приобретают строение фиброцитов, а хрящ – строение сухожилия. С другой стороны волокнистый хрящ постепенно переходит в гиалиновый. Волокнистая хрящевая ткань отличается повышенной механической прочностью. Костные ткани Костные ткани состоят из клеток и минерализованного межклеточного вещества (67% - минеральные компоненты; 33% - органические). Клетки костной ткани Остеобласты – молодые, функционально активные клетки. В зрелой кости местами их локализации являются: · надкостница; · эндост; · каналы остеонов. В формирующейся костной ткани остеобласты располагаются на поверхности костного матрикса. Активные остеобласты имеют кубическую форму, тонкие отростки, связывающие их с другими клетками, светлое ядро, базофильную цитоплазму, хорошо развитые органеллы белкового синтеза. Функции остеобластов: · остеобласты синтезируют и секретируют органический компонент. межклеточного вещества (остеоид); Органические компоненты костной ткани: коллаген I типа (90% всех белков), коллагены других типов (III,IV,V,IX,XIII типов) гликопротеины (остеонектин, остеокальцин), протеогликаны; · остеобласты осуществляют процесс минерализации костного матрикса – отложение кристаллов гидроксиапатита и других неорганических солей вдоль фибрилл коллагена - путём синтеза и секреции неколлагеновых белков, контролирующих процесс минерализации, в частности щелочной фосфатазы; · остеобласты участвуют в регуляции потока ионов Са и Р в костную ткань и обратно. · остеобласты синтезируют и выделяют различные ростовые факторы, в том числе и морфогенетические белки кости, определяющие превращение остеогенных клеток в остеобласты. Остеоцит – основной тип клеток зрелой костной ткани. Образуются из остеобластов, когда те в результате своей синтетической активности и минерализации оказываются окруженными со всех сторон минерализованным матриксом. При этом происходит утрата способности к делению, клетки уменьшаются в размерах, их синтетическая активность резко падает. Уплощенные тела остеоцитов лежат в полостях - лакунах, а их отростки – в костных канальцах – узких тоннелях в твёрдом минерализованном матриксе. Своими отростками остеоциты контактируют
друг с другом при помощи щелевых соединений. Благодаря этому создаётся единая сеть взаимодействующих клеток, связанная при помощи структурных гликопротеинов (остеонектин) с межклеточным веществом. Основная функция остеоцитов - поддержание нормального состояния костного матрикса и баланса кальция и фосфора в организме. Данный дифферон включает клетки следующих стадий развития: остеогенные клетки → преостеобласты → остеобласты → остеоциты. Остеокласты имеют иное – гематогенное – происхождение. Остеокласты образуются из моноцитов крови путём их слияния с формированием многоядерных гигантских клеток (точнее, симпластов).
· остеокласты осуществляют разрушение (резорбцию) костной ткани; · поддержание минерального гомеостаза – в связи с высвобождением при разрушении матрикса большого количества минеральных веществ, поступающих в кровь. Остеокласты имеют крупные размеры и большое число - до 20-50 - ядер. Остеокласты располагаются поодиночке в образованных ими углублениях костной ткани (в резорбционных лакунах, лакунах Хаушипа). Цитоплазма – ацидофильная, содержит хорошо развитый комплекс Гольджи, многочисленные лизосомы и мембранные пузырьки, митохондрии. Маркёрные ферменты остеокластов – особая форма кислой фосфатазы, карбоангидраза и АТФаза. Участок цитоплазмы, прилежащий к костному матриксу, образует многочисленные складки клеточной мембраны - гофрированный край. Плотное прикрепление образуется в зоне краевых светлых зон, в результате чего создается герметичность лакуны и зоны резорбции. Закисление содержимого лакун осуществляется: (а) путем выделения кислого содержимого вакуолей; (б) благодаря действию протонных насосов, накачивающих ионы Н+ в лакуны (источник протонов – реакция между СО2 и Н20 при помощи фермента карбоангидразы). Кислое содержимое вызывает резорбцию минерального компонента; выделение лизосомальных ферментов – растворение органических компонентов. Регуляция активности обеспечивается общими и локальными факторами. В частности гормоны тирокальцитонин и женские половые гормоны подавляют функции остеокластов, а гормон паращитовидной железы паратирин стимулирует их активность. Образование костной ткани (остеогистогенез) Источником развития костных тканей является склеротомная мезенхима. Различают два способа развития костной ткани – прямой остеогистогенез и непрямой остеогистогенез. Прямой остеогенез (развитие костной ткани непосредственно из мезенхимы) характерен для развития грубоволокнистой костной ткани, образующей первоначально плоские кости черепа, ключицы и др. Происходит уже в первый месяц эмбриогенеза. Прямой остеогенез включает три основные стадии: · формирование остеогенного островка (путем скопления активно размножающихся клеток мезенхимы в участке развития будущей кости); · дифференцировка клеток остеогенного островка (в остеобласты) и образование остеоида; остеоид состоит из коллагеновых (оссеиновых) волокон, гликозаминогликанов, протеогликанов и гликопротеинов. · обызвествление (минерализация) остеоида; замурованные остеобласты превращаются в остеоциты. Образуются костные балки, трабекулы, которые затем сливаются в единую сеть. · Перестройка грубоволокнистой костной ткани в пластинчатую. СОЕДИНИТЕЛЬНЫЕ ТКАНИ ОБЩИЕ ПРИЗНАКИ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ • развитие в эмбриональном периоде из общего источника – мезенхимы; • высокое содержание межклеточного вещества (которое состоит из волокон и основного аморфного вещества). В некоторых тканях (хрящ, кость) межклеточное вещество играет функционально ведущую роль. ФУНКЦИИ СОЕДИНИТЕЛЬНЫХ ТКАНЕЙ • поддержание постоянства внутренней среды организма – гомеостаза, что включает функции: • трофическая (обеспечивает питательными веществами другие ткани); • регуляторная (влияет на деятельность других тканей посредством биологически активных веществ и контактных взаимодействий); • защитная (иммунные реакции и воспаление протекают в соединительной ткани); • опорная, формообразующая - соединительные ткани образуют: - строму различных органов; - капсулы различных органов, связанных со стромальными элементами; - опорные органы (сухожилия, связки, хрящи, кости). • пластическая – участие в восполнении объёма разрушенной части органов и тканей (регенерация).
|
Последнее изменение этой страницы: 2019-03-22; Просмотров: 470; Нарушение авторского права страницы





 неразветвленные, отрицательно заряженные, гидрофильные полисахаридные молекулы (хондроитинсульфат, дерматан-, кератан-, гепаран-сульфаты и гепарин). За исключением гиалуроновой кислоты (единственный ГАГ, который не содержит сульфатных групп) ковалентно связываются с белками, образуя протеогликаны. ГАГ синтезируются в грЭПС и комплексе Гольджи фибробластов и других клеток и выделяются в межклеточное пространство, где объединяются в агрегаты. Гиалуроновая кислота – очень длинная молекула - до 5000 дисахаридных субъединиц, - через связывающие белки связывает протеогликаны, формируя трёхмерную молекулярную основу матрикса.
неразветвленные, отрицательно заряженные, гидрофильные полисахаридные молекулы (хондроитинсульфат, дерматан-, кератан-, гепаран-сульфаты и гепарин). За исключением гиалуроновой кислоты (единственный ГАГ, который не содержит сульфатных групп) ковалентно связываются с белками, образуя протеогликаны. ГАГ синтезируются в грЭПС и комплексе Гольджи фибробластов и других клеток и выделяются в межклеточное пространство, где объединяются в агрегаты. Гиалуроновая кислота – очень длинная молекула - до 5000 дисахаридных субъединиц, - через связывающие белки связывает протеогликаны, формируя трёхмерную молекулярную основу матрикса.




 Функции остеокластов:
Функции остеокластов: