
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
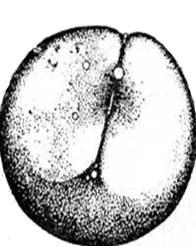
Препарат 5. Гаструла морского ежа (поперечный срез)
На рис. 19 видно, что гаструляция у морского ежа протекает по типу инвагинации с элементами униполярной иммиграции. Бластомеры вегетативного полюса целобластулы несколько крупнее, чем на анимальном полюсе. Часть клеток вегетативного полюса иммигрируют в бластоцель и располагаются там рыхло, формируя первичную мезенхиму. Оставшиеся клетки вегетативного полюса становятся высокими, цилиндрическими и клеточный пласт начинает инвагинировать в бластоцель.
Рис. 19. Последовательные стадии гаструляции у морского ежа (Голиченков, 2006). 1 – бластоцель; Занятие 6 Развитие хрящевых ганоидов (осетровых рыб) Хрящевым ганоидам свойственен тот же примитивный тип развития, какой характерен для амфибий, а также круглоротых, костных ганоидов, кистеперых и двоякодышащих рыб. У осетровых рыб умеренно телолецитальные яйца, полное неравномерное дробление, непрямое развитие с личиночной стадией и метаморфозом. Однако, яйца хрящевых ганоидов более дифференцированы в отношении разделения желточного запаса и образовательной цитоплазмы. Неравномерность в дроблении проявляется в том, что некоторые борозды лишь врезаются в яйцо, не достигая вегетативного полюса, а другие делят яйцо полностью. В результате формируется амфибластула. Мелкие анимальные бластомеры занимают сначала 1/3 поверхности яйца. Гаструляция похожа на таковую амфибий и проявляется в инвагинации и эпиболии. При инвагинации мелкие клетки анимального полушария проникают внутрь через дорзальную и латеральные губы бластопора и формируют крышу архентерона. Здесь располагаются зачатки хорды и мезодермы. Клетки, проходящие внутрь через вентральную губу бластопора, образуют дно архентерона с энтодермальной подушкой. Эпиболия проявляется в наползании микромеров на вегетативные макромеры. Когда микромеры покрывают еще только половину поверхности яйца, инвагинация уже заканчивается, и эти микромеры, лежащие у дорзальной губы бластопора, являются материалом для образования нервной пластинки. Вслед за окончанием гаструляции этот материал образует два сближенных между собой медуллярных валика. К концу обрастания желточной пробки, передний конец медуллярных валиков раздвигается, образуя передний и задний мозговые пузыри. В то же время нервные валики в задней части удлиняются и задним краем нарастают на желточную пробку. К этому моменту начинается сегментация сомитов, в передних из них начинают формироваться пронефросы. После закрытия желточной пробки формирование туловищных сомитов завершается, а вольфовы протоки достигают полной длины. В головной части формируются мозговые пузыри, которые значительно выступают над поверхностью яйца, две пары жаберных щелей (их число затем возрастает до пяти пар), слуховые пузырьки. На заднем конце зародыша в виде бугорка появляется хвостовая почка. Все эти образования распластаны на сферической поверхности яйца. Затем, по мере роста и дифференцировки сомитов, головной отдел зародыша и передние 20 сегментов туловища приподнимаются над поверхностью яйца (желтка). Хвостовая почка увеличивается в длину. На поверхности желтка под передним концом головы образуется сердце. По мере потребления желтка энтодермальной подушки его размеры уменьшаются только в дорзовентральном направлении. Снизу головного отдела и в области 40-го сомита, путем образования наружных выемок и полостей возникают ротовое и анальное отверстия. Происхождение последнего никак не связано с бластопором, который замкнулся на уровне 20-го сомита. В образовании хвостовой почки принимают участие продолжения зачатков всех осевых органов, собирающихся в области бластопора, превратившегося при замыкании нервной трубки в нервно-кишечный канал. При продолжении роста хвостовой почки и достижения зародышем предельной длины, свойственной данному виду, рост зародыша в длину прекращается и нервно-кишечный канал зарастает. Из яйца осетровых рыб развивается личинка, которая имеет внутренние жабры, торчащие из-под жаберной крышки, короткую голову, обонятельные ямки и недоразвитые глаза. По краям рта возникают мелкие острые зубы, сходные по строению с зубами акул. Есть зачатки наружных жабр и брызгальца. Тело личинки сверху и снизу до конца желточного мешка окаймлено плавательной эктодермальной складкой. Личинка имеет зачатки грудных плавников в виде небольших полукруглых складок. Метаморфоз протекает быстро. Образуются жаберные крышки, вырастает рострум. Размеры желточного мешка значительно сокращаются. Формируются спинные, грудные, брюшные и хвостовой плавники. Тело малька покрывается чешуёй (жучками) и появляются накладные кости черепа. Как и у амфибий, метаморфоз у хрящевых ганоидов связан с активностью щитовидной железы.
З Рис. 20. Развитие хрящевых ганоидов (Иванов, 1945). А - начало дробления; Б - бластула; В - начало гаструляции; Г - конец гаструляции; Д - нейрула; Е - обрастание желтка зародышем; Ж - отделение зародыша от желтка; З - личинка Занятие 7 Развитие птиц Птицы являются представителями высших позвоночных, эмбриогенез которых приспособлен к наземным условиям. С развитием птиц лучше всего знакомиться на примере домашней курицы – классическом объекте изучения закономерностей эмбрионального развития. Яйцеклетки птиц – полилецитальные. Зародышевая плазма занимает небольшой участок на анимальном полюсе и представлена зародышевым диском, резко отграниченным от желтка. Собственно яйцеклеткой является то, что в общежитии называют «желтком». В виде голого желтка она выходит из яичника. Сразу же по выходе, в самых начальных частях яйцевода происходит полиспермное оплодотворение с образованием желточной оболочки. Проходя по яйцеводу к клоаке, оплодотворенное яйцо окружается в различных отделах яйцевода белком. В конечном отделе яйцевода образуется скорлупа. Продолжительность развития куриного эмбриона – 21 день. Дробление в курином яйце начинается сразу после оплодотворения, протекает в яйцеводе и продолжается по мере того, как яйцеклетка одевается белковой оболочкой и скорлупой. После откладки яйца до насиживания дальнейшего развития не происходит. Возобновляются процессы развития с момента насиживания. Дробление меробластическое, дискоидальное. Дискобластула представляет собой многоклеточную пластинку, распластанную на желтке, причем между зародышевым диском и желтком обычно имеется небольшая полость – бластоцель. Гаструляция состоит из двух фаз и происходит еще в то время, когда яйцо находится в половых путях самки. I фаза - расслоение (деляминация) зародышевого диска на эпибласт и гипобласт. Впоследствии из первого образуются зародышевые листки и большая часть внезародышевого материала, а из второго – часть внезародышевой энтодермы. Кроме того, в начале инкубации в центре зародышевого диска (эпибласта) выделяется зародышевый щиток. Окружающая его часть зародышевого диска оказывается к этому времени несколько приподнятой над желтком. Эта часть носит название светлого поля - area pellucida (клетки будущего зародыша). Светлое поле окружено плотно прилежащей к желтку бластодермой, получившей название темного поля - area opaca (клетки будущих внезародышевых, провизорных органов: амниона, серозы, аллантоиса). Наружный край темного поля (гипобласта) является краем обрастания. II фаза гаструляции заключается в активных клеточных перемещениях в эпибласте, приводящих к образованию всех трех зародышевых листков как в зародышевой, так и во внезародышевых частях, которые происходят в первые сутки инкубации. Главнейшие перемещения совершаются в области зародышевого щитка. Перемещение клеточного материала здесь идет в направлении от передного к заднему концу будущего зародыша. Клеточные потоки особенно быстро движутся по краям зародышевого щитка и быстрее достигают заднего его края. Два клеточных потока, встречаясь по срединной линии, направляются вперед. Клеточный материал медиальной части движется медленнее. Боковые клеточные потоки в срединной части образуют утолщенную клеточную полоску, которая постепенно удлиняется от задней части зародышевого щитка к передней. Это образование получило название первичной полоски. В месте встречи клеточных потоков, движущихся с переднего и заднего концов зародышевого щитка, образуется бугорок, возвышающийся над зародышевым щитком - первичный или гензеновский узелок. На вершине его появляется углубление - первичная ямка, а по середине первичной полоски возникает первичная бороздка, являющаяся продолжением первичной ямки. Передний край первичной ямки является дорзальной губой бластопора, а края первичной бороздки - латеральными губами. Затем передний край первичной ямки надвигается на лежащий кзади клеточный материал так, что бластопор начинает замыкаться спереди назад и находящийся непосредственно перед первичным узелком клеточный материал подворачивается через дорзальную губу бластопора, формируя между наружным и внутренним листками хордальный или головной отросток. Одновременно, материал первичной полоски начинает погружаться внутрь через латеральные губы бластопора. Он проникает между наружным и внутренним зародышевыми листками, ложась по бокам хордального отростка, распространяясь в стороны и вперед. После гаструляции снаружи остаются только материал нервной пластинки и кожная мезодерма. Участки мезодермальных выростов, расположенные в передних частях первичной полоски, лежащие по сторонам от хордального выроста, дают сомиты. Мезодермальный материал, лежащий в задних частях первичной полоски, смещается вперед позже и входит в состав спланхнотомов. Таким образом, возникают два мезодермальных крыла, свободные края которых разрастаются за пределы зародышевого щитка и вклиниваются между экто- и энтодермой зародышевого диска, продолжаясь во внезародышевую часть. Материал, лежащий снаружи впереди и по бокам от гензеновского узелка, параллельно с образованием хордального отростка, также вытягивается по осевой линии в длину и дает начало нервной пластинке. Формирование зачатков органов и отделение клеточного материала сходно с таковым у ранее изученных групп позвоночных животных. Энтодерма первоначально представлена плоской пластинкой клеток, распластанной над заполненной жидкостью подзародышевой полостью. Лишь позднее зародыш начинает приподниматься над желтком, энтодермальная пластинка выгибается кверху, образуя под хордой кишечный желобок, который позже сворачивается в кишечную трубку. Образование ее начинается на переднем, а затем на заднем конце тела и постепенно распространяется от обоих концов к середине. Как и у хрящевых ганоидов, у птиц формируется желточный мешок, выполняющий функцию переваривания и всасывания желтка, и служащий на ранних этапах развития кроветворным органом и органом дыхания. Нервная пластинка образует желобок, а затем свертывается в трубку, погружаясь под кожную эктодерму. Мезодерма дифференцируется на сомиты, занимающие наиболее осевое положение по бокам хорды и нервной трубки, а также, лежащие латеральнее - нефротомы. Наиболее латерально расположенные участки мезодермы - спланхнотомы. Сомиты и нефротомы представляют сегментированную часть мезодермы, причем сегментация распространяется спереди назад. На более поздних стадиях развития сомиты подразделяются на дерматомы, миотомы и склеротомы, из которых развиваются, соответственно, кориум кожи, мышцы и дефинитивный осевой скелет в виде материала позвоночника, расположенного вокруг хорды. Одновременно, спланхнотомы, расщепившись на париетальный (наружный) и висцеральный (внутренний) листки, образуют целом. Нефротомы дают начало вольфову протоку и канальцам первичной почки. Тело зародыша отделяется от внезародышевой части туловищной складкой. Внезародышевая часть разделена на две пластинки экзоцеломом. Наружная пластинка состоит из эктодермы и париетальной мезодермы и поднимается над телом зародыша в виде амниотической складки. Внутренняя пластинка состоит из висцеральной мезодермы и энтодермы, образуя стенку желточного мешка. В мезодерме видны кровеносные сосуды. Препарат 1. Первичная полоска, тотальный препарат На препарате видно, что зародышевый диск разделился на однослойный гипобласт и многослойный эпибласт (рис. 20). В центральной утолщенной части зародышевого диска (первичная полоска) происходит миграция клеточного материала, который формирует рыхло лежащие мезенхимные клетки (мезодерма) по бокам от первичной полоски.
Рис. 20. Зародыш курицы. 1 - первичная полоска Препарат 2. Первичная бороздка на стадии закладки мезодермы, На препарате видно, что эктодерма и энтодерма зародышевого диска светлой области area pellucida и темной области area opaca отличаются гистологически (рис. 21). Так, клетки экто- и энтодермы во внезародышевой части имеют округлую форму и вакуолизированную цитоплазму. В центральной части зародышевого диска формируется желобок, который и представляет собой первичную бороздку. Под ним располагается материал хордомезодермы, а по бокам располагается мезодерма сомитов и спланхнотомов, которая обнаруживает уже более или менее упорядоченное строение.
Рис. 21. Поперечный разрез куриного зародыша на стадии первичной полоски (17 ч. инкубации).
Рис. 22. Поперечный разрез куриного зародыша (24 часа инкубации). |
Последнее изменение этой страницы: 2019-04-09; Просмотров: 170; Нарушение авторского права страницы

 А
А  Б
Б В
В  Г
Г Д
Д  Е
Е Ж
Ж


