|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Второе пришествие трихроматических обезьян
Наличие полноценного цветового зрения у ревунов, обнаруженное во время масштабного исследования обезьян Нового Света, оказалось для ученых неожиданностью. Эта способность у ревунов, как и у приматов Старого Света, могла возникнуть в результате реализации одного из двух альтернативных эволюционных сценариев. В первом случае ревуны могли унаследовать цветовое зрение от своего общего предшественника с обезьянами Старого Света. Во втором, который, как я уже говорил, и имел место, ревуны должны были обзавестись цветовым зрением в результате последовательности событий, не связанной с эволюцией обезьян Старого Света. Откуда мы знаем, какой сценарий правильный? Допустим, какие-либо два вида организмов имеют общие признаки. Чтобы установить, унаследованы ли они от общего предшественника или появились независимо, нужно посмотреть, как связаны эти виды на эволюционном древе. Отмечая наличие или отсутствие признака на эволюционном древе, можно проследить историю возникновения этого признака. Каждая точка ветвления на древе соответствует общему предку исходящих из этой точки ветвей. Если у всех видов, произошедших от данного предка, этот признак есть, то, скорее всего, предок также обладал этим признаком (рис. 6.1 а). Однако если две ветви, обладающие данным признаком, соединяются с другими ветвями, в которых этого признака нет, то, вероятнее всего, в двух ветвях этот признак появился независимым образом (рис. 6.1 б).
Рис. 6.1. Альтернативные варианты появления общих признаков у разных видов. Разные виды организмов могут обладать общим признаком (звездочка), если они унаследовали его от общего предка (A) или в результате независимой эволюции этого признака у данных видов после их отделения от общего предка (Б). Рисунок Лианн Олдс.
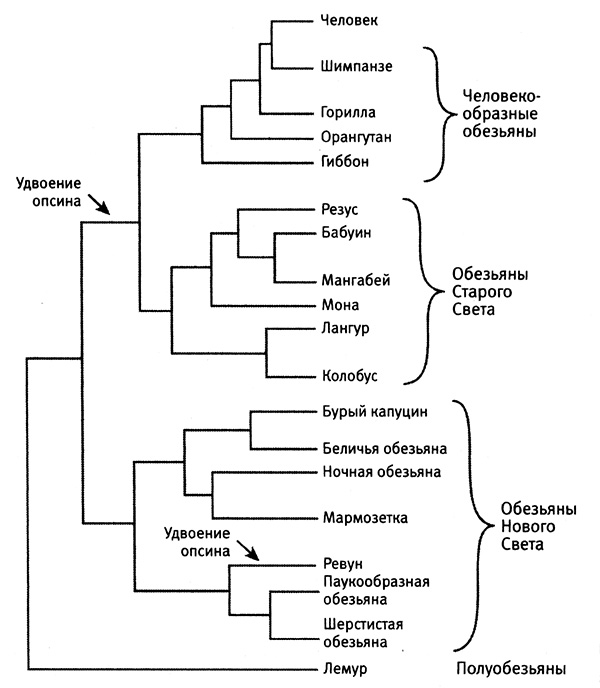
Теперь давайте взглянем на эволюционное древо приматов Старого и Нового Света (построенное на основании наследования SINE, LINE и других последовательностей ДНК), изображенное на рис. 6.2.
Рис. 6.2. Полноценное цветовое зрение возникало у приматов дважды. На основании распределения способности к цветовому восприятию и эволюционных связей между приматами, представленных на этой диаграмме, можно сделать вывод о том, что полноценное цветовое зрение возникало у приматов дважды (стрелки): один раз у общего предка человекообразных и других обезьян Старого Света, второй раз — у ревунов. Из статьи Giladet at., 2004, PloSBiology 2 (1): e5 (с изм.).
Из рисунка следует, что ближайшими родственниками ревунов являются другие обезьяны Нового Света, но ни один из этих видов не обладает полноценным цветовым зрением. Теоретически возможно, что у предшественника обезьян Нового Света цветовое зрение было, но все потомки, кроме ревуна, эту способность потеряли. Однако такой сценарий предполагает лишь одно приобретение и слишком много потерь. Более простое объяснение, подразумевающее гораздо меньшее количество эволюционных изменений, заключается в том, что ревун произошел от предшественника с дихроматическим зрением и самостоятельно приобрел способность различать цвета. К счастью, чтобы докопаться до истины, мы не обязаны полагаться лишь на информацию, заключенную в эволюционном древе. Удвоение генов оставляет в ДНК свои следы. Анализ последовательности и организации современных генов опсинов в ДНК приматов Старого Света, а также ревунов и других обезьян Нового Света проливает свет на их историю. Из текста ДНК однозначно следует, что удвоение гена опсина у приматов Старого Света и у ревуна — несвязанные события. Мы можем это утверждать, поскольку в этих двух случаях размеры удвоенных фрагментов ДНК разные. У приматов Старого Света вне кодирующей последовательности каждого из двух генов имеются одинаковые последовательности длиной по 236 пар оснований. Это означает, что при создании гена средневолнового опсина была удвоена еще и прилежащая последовательность из 236 пар оснований. А у ревуна к удвоенным генам опсина примыкают гораздо более протяженные некодирующие участки. Такое могло произойти лишь в том случае, если удвоение гена опсина у ревуна произошло независимо от удвоения гена опсина у нашего с вами предка. Кроме того, дополнительным доказательством независимости двух событий является степень сходства генов в каждой паре. Со временем после удвоения гена в каждой его копии накапливаются мутации. Чем раньше произошло удвоение, тем больше различие между двумя копиями. Так вот, у приматов Старого Света последовательности двух генов опсина различаются более чем на 5 %, тогда как гены ревуна различаются лишь на 2,7 %. Это означает, что удвоение гена опсина у ревуна произошло позднее, чем у приматов Старого Света. Это заключение вполне согласуется с палеонтологическими доказательствами более поздней эволюции обезьян Нового Света, начавшейся после разделения южноамериканского и африканского континентов. Конвергентная эволюция опсинов ревуна и приматов Старого Света не сводится лишь к удвоению генов. Вспомним, что для полноценного цветового восприятия необходима тонкая настройка опсинов на разные длины волн и что на значение максимума поглощения опсинов MWS и LWS влияют аминокислоты, расположенные в трех ключевых позициях этих молекул. Средне- и длинноволновые опсины ревуна настроены на те же самые длины волн, а в трех ключевых позициях имеют те же самые аминокислотные остатки, что и аналогичные опсины человека и других приматов Старого Света. Это означает, что в ходе эволюции «нового» опсина MWS у ревуна и «нового» опсина MWS у приматов Старого Света произошло три одинаковые замены. Анализ ДНК свидетельствует о том, что эволюция зрения и обоняния у ревунов шла тем же путем, что и у приматов Старого Света за миллионы лет до этого. Удвоение гена опсина, тонкая настройка ключевых позиций в молекулах опсинов и отмирание генов, ответственных за обоняние, повторились в той же последовательности и где-то с теми же деталями. Изучение конвергентной эволюции цветового зрения у приматов и восприятия ультрафиолетового света у некоторых птиц показывает, что родственные виды могут приобретать похожие признаки, однако конвергенция ни в коем случае не ограничена степенью родства. Вспомните историю эволюции родопсина, настроенного на восприятие тусклого синего света, у глубоководного угря и у афалины (глава 4), а именно появление трех идентичных аминокислотных остатков в ключевых положениях их опсинов. Одна и та же история, но разные гены и крайне далекие друг от друга виды. История эволюции опсинов вызывает вопрос более общего плана: насколько часто появление одинаковых признаков у разных видов организмов связано со сходными эволюционными изменениями ДНК? Давайте рассмотрим четыре примера того, как с помощью похожих механизмов с участием одного и того же гена или одних и тех же генов у разных видов и в разное время возникали одинаковые признаки.
Одним путем к одной цели
Задолго до появления у обезьяны колобуса особой пищеварительной системы и способности переваривать листья это же свойство развилось у предка таких обычных жвачных животных, как коровы, овцы и козы. Есть ли какое-то сходство в эволюции пищеварения у обезьян и у коров? Безусловно, есть. Вспомним, что одним из механизмов адаптации к новому способу питания у колобуса было появление специализированной панкреатической рибонуклеазы, расщепляющей питательные вещества в смеси листьев и бактерий. Этот фермент возник в результате удвоения и тонкой настройки гена, кодирующего обычную рибонуклеазу. У коровы этот ген также удвоился и подвергся тонкой настройке в соответствии с условиями в пищеварительной системе этого животного. Мы знаем, что эти события у обезьян и коров произошли независимо, поскольку удвоение гена рибонуклеазы произошло у всех жвачных, но у ближайших родственников жвачных, таких как гиппопотамы и дельфины, а также у ближайших родственников колобуса ген рибонуклеазы только один. Таким образом, эти группы жвачных животных не могли унаследовать удвоенные гены от общего предшественника. Кроме того, африканские колобусы — не единственные жвачные обезьяны. В Азии существует еще одна группа жвачных обезьян. Эффектный немейский тонкотел (рис. L цветной вкладки) — находящийся под угрозой вымирания вид обезьян, распространенный во Вьетнаме, Лаосе, Камбодже и Китае, — также имеет удвоенный ген рибонуклеазы.
Жианши Цанг из Университета Мичигана обнаружил, что удвоение гена рибонуклеазы у обезьян происходило в разное время и приводило к образованию разного количества генов (трех у африканских видов, двух у азиатских). Однако впоследствии в ферментах произошло несколько совершенно идентичных изменений. Вероятность случайного совпадения изменений у двух групп обезьян чрезвычайно низка. Гораздо более вероятно, что параллельные изменения в структуре ферментов у обеих групп обезьян являются результатом естественного отбора, настраивавшего ферменты на работу в более кислой среде передней кишки. Возникновение ископаемых генов и полное исчезновение генов тоже повторяются. В предыдущей главе я рассказывал об избирательной потере функций семи генов метаболизма галактозы у дрожжей S. kudriavzevii. Но существуют еще три вида дрожжей, принадлежащие к разным родам и разделенные миллионами лет эволюции, которые также потеряли все или почти все гены, связанные с метаболизмом галактозы, и больше не могут расщеплять этот сахар. На основании эволюционного родства между дрожжами можно с уверенностью утверждать, что в ходе эволюции эта группа генов независимым образом была потеряна как минимум три раза, а может быть, и больше. Скорее всего, в каждом случае ослабевало влияние естественного отбора, что и приводило к разрушению и исчезновению генов. Ослабление влияния естественного отбора также объясняет повторяющуюся эволюцию признаков у животных, обитающих в пещерах. Например, известно множество видов пещерных рыб, которые лишились глаз и окраски. Поскольку эти рыбы относятся к разным семействам, куда также входят рыбы, живущие в поверхностных водах и обладающие зрением, становится ясно, что потеря зрения и пигментации происходила в истории многократно. Пещерные рыбы позволяют выяснить, имеют ли эти внешние признаки какие-то более глубокие общие основания. Не так давно Мередит Протас и Клифф Табин с медицинского факультета Гарвардского университета, Билл Джеффри из Университета Мэриленда и их сотрудники изучили эволюцию альбинизма у слепых рыб, обитающих в поземных водоемах в мексиканских пещерах (рис. 6.3).
Рис. 6.3. Эволюция альбинизма у слепых пещерных рыб. В то время как живущие в поверхностных водах представители вида Astyanax mexicanus выглядят как обычные рыбы, многие их пещерные родственники, например обитатели пещер Молино и Пачон, в ходе эволюции не раз теряли зрение и окраску в результате мутации в одном и том же гене. Фотография любезно предоставлена Мередит Протас и Клиффом Табином из Гарвардского университета.
Эти рыбы (Astyanax mexicanus ) принадлежат к тому же отряду, что и пираньи и ярко окрашенный неон-тетра, однако обитатели примерно 30 пещер на территории Мексики потеряли свою окраску. Ученые обнаружили, что в двух исследованных ими популяциях рыб в результате делеции фрагмента ДНК был инактивирован один и тот же ген, ответственный за пигментацию, однако делеции в каждой популяции были разными. Это доказывает, что популяции, населяющие разные пещеры, утратили окраску независимым образом. Развитие альбинизма у пещерных рыб проще всего объяснить ослаблением естественного отбора. Какая разница, как вы выглядите, если вокруг темно? Однако для многих других животных окраска тела важна для выбора партнера, спасения от врагов и других важных функций, находящихся под влиянием естественного или полового отбора. Один из самых распространенных вариантов окраски — черный. У многих видов мех, чешуя или перья в той или иной степени окрашены в черный цвет. Существуют и вариации внутри одного вида — особи разного пола или из разных популяций могут различаться по количеству черных полос или пятен на теле. Во многих случаях естественный или половой отбор влиял на один и тот же ген, ответственный за окраску тела позвоночных животных. Например, голубые северные (снежные) гуси бывают либо белыми, либо «голубыми». «Голубой» цвет связан с появлением в перьях черного пигмента (рисунки M и N цветной вкладки).
Окраска гусей меняется в зависимости от места их обитания: голубые гуси чаще встречаются на востоке Канады, а белые — в самой западной части ареала, в Восточной Сибири. Различие в окраске имеет значение при выборе полового партнера. Молодые гуси очень рано запоминают цвет перьев своих родителей и впоследствии предпочитают спариваться с особями того же цвета. За вариации окраски у северных гусей отвечает одно-единственное генетическое различие. Все дело в гене рецептора меланокортина-1 (MC1R ). Различие между генами MC1R у белых и голубых гусей состоит всего в одном триплете, кодирующем аминокислоту в позиции 85. У других птиц вариации последовательности гена MC1R также коррелируют с изменением окраски. Темный и желтый варианты окраски бананового певуна (сахарной птицы) определяются единственным основанием в гене MC1R. Однако эта единственная замена в данном случае происходит в ином положении, чем у голубых и белых северных гусей. Замена еще в одной позиции в данном гене определяет наличие светлого или темного оперения у третьего вида птиц — короткохвостого поморника (рис. O цветной вкладки).
У этих птиц окраска оперения также играет определяющую роль в выборе партнера и находится под влиянием полового отбора. Необыкновенные различия в окраске самцов и самок прекрасных расписных малюров также определяются геном MC1R (рис. цветной вкладки P и Q ).
Однако роль гена MC1R в эволюции окраски тела не ограничена миром птиц. Вариации MC1R ответственны за различия между оранжевыми и черными ягуарами, белыми и темными представителями бурых медведей на западе Северной Америки, светлыми и темными ящерицами, а также за варианты окраса домашних собак, кошек и лошадей. Одним из наиболее изученных примеров влияния гена MC1R на эволюцию диких животных является изменение окраски мешотчатых прыгунов, распространенных в пустынных юго-западных районах США. Во второй главе на примере генетики и эволюции светлых и темных вариантов прыгунов я проиллюстрировал связь между случайными мутациями, отбором и временем. Эти грызуны обитают в песчаных пустынях и на покрытых окаменелой черной лавой участках земли в Аризоне и Нью-Мексико. Соответствующая окраска позволяет этим животным быть менее заметными на фоне светлого песка или черной лавы. Майкл Нахман, Хопи Хекстра и их коллеги из Университета Аризоны показали, что живущие в районе пустыни Пинакате темные прыгуны отличаются от светлых прыгунов четырьмя позициями в белке MC1R. Интересно, что темные прыгуны имеют точно такую же замену в положении 230, что и короткохвостый поморник. Таким образом, мало того что один и тот же ген задействован в эволюции окраски у некоторых видов птиц, рептилий и млекопитающих, в некоторых случаях у разных видов в этом гене произошли совершенно одинаковые замены. Еще один пример точного повтора эволюционных изменений — изменение окраски ягуарунди и золотистоголовой львиной игрунки. У темноокрашенных ягуарунди в гене MC1R обнаружена делеция 24 оснований. Точно такая же делеция присутствует у золотистоголовых игрунок, у которых, в отличие от других львиных игрунок, все остальное тело черное (цветная вкладка R ).
Эволюция жвачных млекопитающих, метаболизм галактозы у дрожжей, альбинизм у пещерных рыб и темная окраска тела у различных птиц, рептилий и млекопитающих — все это иллюстрирует повторяемость эволюционных изменений на фундаментальном уровне, то есть на уровне отдельных генов. В ранее приведенных примерах конвергентной эволюции опсинов это воспроизведение было настолько точным, что иногда затрагивало одну и ту же пару оснований. В тех случаях, о которых я упомянул только что, изменения воспроизводятся не с такой высокой точностью. Подробное изучение биохимических свойств рибонуклеазы и рецептора MC1R показало, что в каждом из этих белков есть множество различных участков, изменение которых приводит к похожим результатам. Разница между идентичным воспроизведением эволюции опсинов и не столь точным повторением эволюции других белков показывает, что для одних «проблем» (или адаптаций) эволюция находит множество решений, а для других — лишь одно. Структура опсина такова, что лишь наличие правильных аминокислотных остатков в нескольких ключевых позициях обеспечивает настройку пигмента на восприятие определенной длины волны. Структуру и активность рибонуклеазы и MC1R настроить легче, поэтому существует много разных способов их изменения. Другими словами, в случае некоторых генов и некоторых признаков для получения одного и того же биологического эффекта генетический текст не обязательно должен меняться одинаково. Более того, оказывается, что в некоторых случаях конвергентная эволюция может начаться из совершенно разных генетических исходных точек.
|
Последнее изменение этой страницы: 2019-05-08; Просмотров: 54; Нарушение авторского права страницы