|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Становление обмена веществ
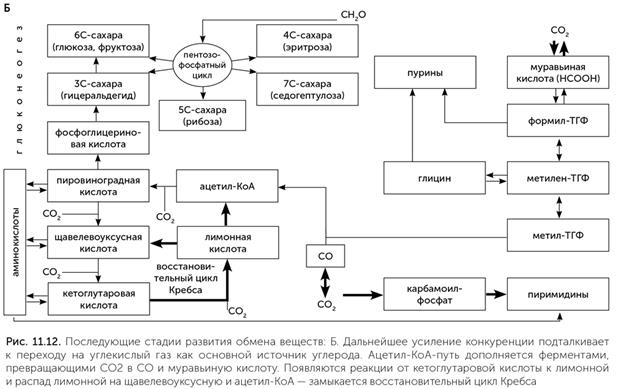
Современный обмен веществ появился не сразу. Скорее всего, его становление прошло через много последовательных стадий. Мы можем попробовать восстановить путь развития обмена веществ начиная от мира РНК и до клеточных организмов современного типа, таких как бактерии. Обмен веществ должен делать нужные для организма вещества из тех, что есть в наличии. Для организмов РНК-мира были нужны прежде всего нуклеотиды и их компоненты – рибоза и азотистые основания. Круг веществ, которые были доступны для РНК-организмов во внешней среде, мы уже очертили в главах 6 и 7: это углекислый газ, формальдегид, муравьиная кислота, формамид, синильная кислота и ее производные (цианамид и цианоацетилен) и, скорее всего, аммиак (NH3) и угарный газ (СО). Для получения рибозы из этого списка лучше всего подходит формальдегид, легко вступающий в реакцию Бутлерова. В современных клетках взаимные превращения сахаров (в цикле Кальвина и пентозофосфатном цикле) происходят по тому же механизму альдольной конденсации, что и стадии реакции Бутлерова. Более того, у ряда метилотрофных микробов мы встречаем рибулозо-монофосфатный цикл, производящий сахара непосредственно из формальдегида. Скорее всего, у РНК-организмов действовал какой-то примитивный вариант рибулозо-монофосфатного цикла под контролем рибозимов. Азотистые основания нуждаются в других веществах-предшественниках. До появления жизни они могли строиться из цианида или формамида, как было рассказано в главе 7. В клетках для построения азотистых оснований используются аминокислоты (глицин и аспарагиновая кислота), муравьиная кислота (в форме формил-ТГФ или формил-фосфата), углекислый газ и карбамоил-фосфат. Кроме того, часть азота для пуриновых оснований поступает в составе третьей аминокислоты – глутамина, но его можно заменить просто достаточным количеством аммиака. Муравьиная кислота, СО2 и аммиак доступны в среде. Карбамоил-фосфат похож на формамид, соединенный с фосфатной группой, и возможно, занял место формамида в древнейшем обмене веществ. Глицин образуется в глициновом цикле, например, из муравьиной кислоты и аммиака. Сложнее определить источник аспарагиновой кислоты. В клетках она образуется из щавелевоуксусной кислоты и аммиака. Щавелевоуксусная кислота является промежуточным продуктом цикла Кребса. Щавелевоуксусная и другие кислоты цикла Кребса образуются, например, из СО2 на кристаллах сульфида цинка при освещении, или из формамида на кристаллах оксида титана, или из синильной кислоты при чередовании освещения и нагревания в темноте. Иными словами, в среде, где есть кристаллы сульфида цинка, формамид или синильная кислота, какое-то количество аспарагиновой кислоты неизбежно получится. РНК-организмы могут при помощи рибозимов ускорять и стабилизировать любой из этих путей получения аспарагиновой кислоты. Иначе говоря, на этой стадии обмен веществ РНК-организмов имеет два относительно независимых блока: сахарный (родственный реакции Бутлерова и рибулозо-монофосфатному циклу), питаемый формальдегидом, и одноуглеродный, питаемый муравьиной кислотой и производящий глицин и пуриновые нуклеотиды (рис. 11.11. А). Связь между этими двумя блоками осуществлялась через формальдегид и метилен-ТГФ. Метилен-ТГФ может распадаться, давая формальдегид, если во внешней среде его вдруг окажется недостаточно для синтеза сахаров. Кроме того, есть какие-то химические реакции для получения аспарагиновой кислоты, но мы не знаем, какие именно. Переход ко второй стадии связан с появлением первых белков и увеличением потребности в разнообразных аминокислотах для их построения. Большинство аминокислот, входящих в состав белков, образуются из трех кислот цикла Кребса: пировиноградной, щавелевоуксусной и кетоглутаровой. Все эти кислоты образуются из формамида или на кристаллах сульфида цинка, но с небольшим выходом, поэтому с переходом к белкам нужен более надежный их источник. К этому моменту РНК-организмы уже освоили синтез сахаров. Сахара – не очень стабильные вещества, которые постепенно разлагаются, особенно при повышенной температуре. Основным продуктом их распада является пировиноградная кислота. Различные сахарофосфаты (глюкозо-6-фосфат, рибозо-5-фосфат, глицеральдегид-3-фосфат) превращаются в пировиноградную кислоту просто при нагревании до 70 °C в растворе с солями железа (Keller, 2014). Превращение глюкозы в пировиноградную кислоту в три шага (через глюконовую кислоту и кетодезоксиглюконовую кислоту) происходит с достаточной скоростью даже при комнатной температуре в присутствии двух аминокислот – цистеина и гистидина (Shimizu et al., 2008). Эти и другие подобные реакции, постепенно переходя под контроль рибозимов и ферментов, были примитивными предшественниками гликолиза. Превращение пировиноградной кислоты в щавелевоуксусную и далее в кетоглутаровую происходит в восстановительном цикле Кребса. Эти реакции проходят на кристаллах сульфида цинка, но с небольшой эффективностью, и с повышением потребности в аминокислотах были взяты под контроль рибозимов и ферментов. На этой стадии возникает примитивный гликолиз и часть восстановительного цикла Кребса – от пировиноградной кислоты до кетоглутаровой (рис. 11.11. Б). По мере развития жизни конкуренция за основные ресурсы – муравьиную кислоту и формальдегид – обостряется. В выигрыше оказываются те организмы, которые сумели освоить новые источники углерода. Например, это мог быть угарный газ (СО). Третья стадия развития обмена веществ начинается с появления фермента ацетил-КоА-синтетазы. Этот фермент соединяет метильную (СН3) группу метил-ТГФ с молекулой угарного газа и коферментом А, образуя ацетил-КоА. Появление ацетил-КоА-синтетазы открывает доступ к новому источнику углерода – угарному газу и позволяет пережить нехватку муравьиной кислоты. Вторым следствием появления этого фермента становится возникновение новой связи между одноуглеродным блоком обмена веществ, построенным вокруг реакций с участием витамина В9, и остальным обменом веществ (рис. 11.12. А). Если раньше для превращения муравьиной кислоты в рибозу и аминокислоты приходилось использовать ядовитый формальдегид в качестве промежуточного продукта, то теперь появилась возможность заменить этот путь на более безопасный, через ацетил-КоА и пировиноградную кислоту. Если раньше пировиноградная кислота получалась из сахаров, то теперь, наоборот, нужен способ получать сахара из пировиноградной кислоты. Для получения рибозы и других сахаров из ацетил-КоА через пировиноградную кислоту возникает глюконеогенез – обращение реакций гликолиза. Так путь от доступных в окружающей среде соединений углерода (муравьиная кислота и СО) к рибозе стал намного длиннее, но безопаснее.
Далее в истории метаболизма происходят новые изменения. Протоклетки снижали свою зависимость от сульфида цинка и других минеральных катализаторов, замещая их ферментами, а нарастающий дефицит муравьиной кислоты, формамида и угарного раза вынуждал их переходить к потреблению углекислого газа. Появляется формат-дегидрогеназа, содержащая молибден: она восстанавливает СО2 до муравьиной кислоты для работы фолатного пути. Наконец, у ацетил-КоА-синтетазы возник второй активный центр с никель-железосерным кластером, позволяющий ей восстанавливать СО2 до СО. Так появился полностью автотрофный обмен веществ, способный строить все аминокислоты, нуклеотиды и витамины из углекислого газа в качестве единственного источника углерода, а также обладающий высокой устойчивостью за счет дублирования путей фиксации СО2 (рис. 11.12. Б). Такой обмен веществ, скорее всего, и был у последнего всеобщего предка клеточной жизни.
Итак, обмен веществ развивался и усложнялся в двух направлениях. Во-первых, он переходил от сложных и дефицитных исходных веществ к простым и легкодоступным: от рибозы и азотистых оснований – к формальдегиду и цианиду, потом – к муравьиной кислоте и угарному газу и, наконец, – к углекислому газу. Во-вторых, обмен веществ сначала строился вокруг производства компонентов РНК (рибозы и азотистых оснований), а затем основным направлением стало получение аминокислот для сборки белков. Давний спор о способе питания первых живых организмов (автотрофный или гетеротрофный), похоже, разрешился. Первые РНК-организмы питались готовыми азотистыми основаниями и рибозой, т. е. гетеротрофным способом. Потом они усложнялись и переходили к питанию более простыми веществами. Хотя все эти готовые органические вещества (рибоза, азотистые основания, цианид, формальдегид, муравьиная кислота) в конечном итоге получались из СО2, это происходило без участия живых организмов. В современном мире круговорот углерода замыкается в основном благодаря живым организмам: автотрофы производят органику из СО2, а гетеротрофы потребляют органику, в конечном итоге превращая ее обратно в углекислый газ. Но пока жизнь делала самые первые шаги, она не могла поддерживать все эти процессы, и часть углеродного круговорота, освоенная потом автотрофами, обеспечивалась только силами неживой природы. Какое-то время организмы позднего РНК-мира и раннего РНК-белкового мира сочетали усвоение углекислого газа, муравьиной кислоты и при случае также формальдегида и угарного газа. Иначе говоря, они сочетали в себе признаки автотрофов и гетеротрофов. Здесь хорошо видно, что местами между ними проведена довольно искусственная граница. Муравьиная кислота – органическое вещество, а угарный газ – неорганическое. Поэтому микробы, способные питаться угарным газом, должны называться автотрофами, а потребители муравьиной кислоты – гетеротрофами, хотя различия между ними сводятся буквально к одному-двум ферментам. В конечном итоге жизнь освоила построение всех необходимых молекул из СО2, и появились действительно автотрофные клетки. Но произошло это, судя по всему, уже после появления белков и системы их производства (рибосомы). До этого жизнь успела достичь немалой сложности, питаясь муравьиной кислотой. Такие привычные нам пути обмена веществ, как гликолиз, похоже, появились позднее. Классический гликолиз состоит из 12 последовательных реакций, и все ради того, чтобы выжать немного энергии в виде АТФ из превращения глюкозы в пируват. Если нам просто нужно получить пируват из сахаров, достаточно использовать упрощенный гликолиз, проходящий в три стадии при помощи свободных аминокислот, т. е. даже без ферментов. С выходом клеток из геотермальной «колыбели» с ее разнообразными источниками энергии в большой мир обмен веществ оптимизировался по экономичности. В том числе возник гликолиз современного типа, позволяющий получать энергию путем брожения сахаров.
Глава 12 |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 87; Нарушение авторского права страницы