|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
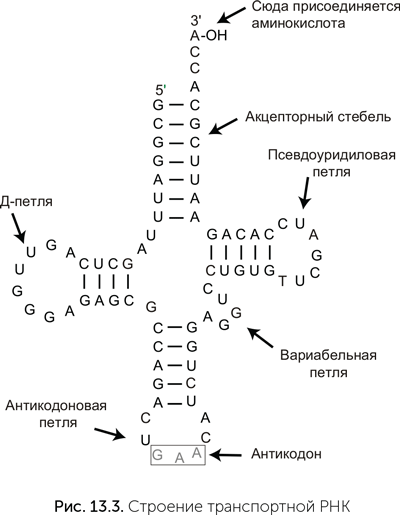
Происхождение транспортных РНК
Транспортные РНК (тРНК) – скорее всего, самая древняя деталь системы синтеза белка. Все эти небольшие (обычно 76 нуклеотидов длиной) молекулы имеют одинаковую пространственную укладку с четырьмя двухспиральными участками, которая на плоскости изображается в виде клеверного листа (рис. 13.3). На «черешке» у всех тРНК есть свободный одноцепочечный фрагмент ССА (цитозин-цитозин-аденин), к которому прикрепляется аминокислотный остаток. Но эти «клеверные листы» используются не только в производстве белков. Как выяснилось, тРНК-подобные структуры участвуют в копировании геномов РНК-вирусов. Такая структура есть, например, на конце одноцепочечного РНК-генома бактериофага[9] Qβ, причем для его узнавания вирусная РНК-полимераза нуждается в помощи бактериальных белков, в норме связывающих транспортные РНК (Weiner and Maizels, 1987). В РНК-вирусах растений встречаются еще более точные аналоги – например, тРНК-подобную структуру из вируса желтой мозаики турнепса узнают бактериальные ферменты, работающие с собственными бактериальными тРНК (Joshi et al., 1984)! Другие вирусы, например, вирус мозаики цветной капусты, используют клеточные тРНК как затравки для синтеза первой цепи ДНК на матрице РНК (Hohn et al., 1985).
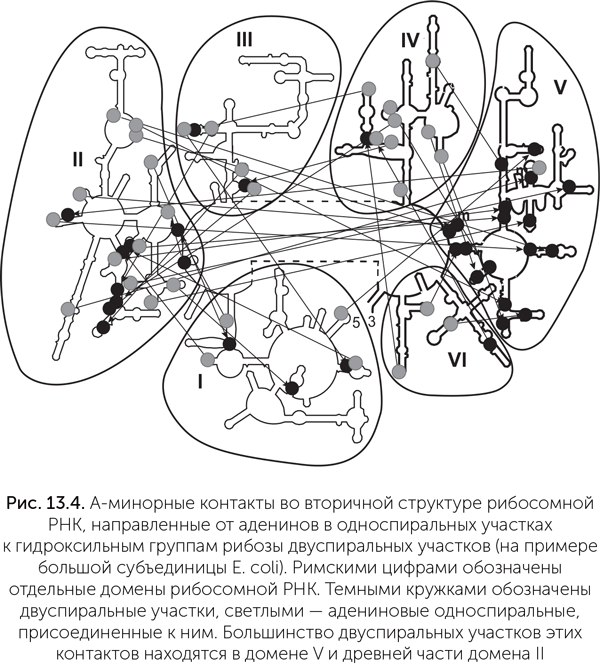
У этих и других вирусов создание новой цепи РНК всегда начинается с двух гуаниновых нуклеотидов, комплементарных двум цитозинам в одноцепочечном ЦЦА-участке тРНК-подобной структуры (рис. 13.2). Гуанин в данном случае выбран, скорее всего, потому, что он образует больше всего водородных связей как с другой цепью РНК, так и с молекулой фермента. Таким образом, аденин на самом конце РНК-матрицы не будет скопирован и должен быть достроен отдельно. И в самом деле, различные вирусные РНК-полимеразы (например, бактериофага Qβ ) и даже клеточные ДНК-полимеразы, доходя до конца матричной цепи, добавляют на конце растущей цепи один лишний аденин. Это особенно странно для бактериальных ДНК-полимераз, которые копируют кольцевые ДНК и в норме никогда не сталкиваются с концом матричной цепи. Еще одна странность проявляется в процессе созревания тРНК: сначала специальный фермент РНКаза Р обрезает концы незрелых транспортных РНК, удаляя в том числе нуклеотиды ССА, а потом ССА достраиваются обратно другим ферментом – нуклеотидилтрансферазой. Из всех этих фактов участия тРНК-подобных структур в репликации была построена гипотеза «геномной метки» (genomic tag hypothesis). Подробное изложение см. в сборнике «RNA World» (Cold Springs Harbor press, 1999). Эта гипотеза утверждает, что в РНК-мире тРНК-подобные «клеверные листы» появились на концах геномных молекул. Они служили местом начала копирования и защитными концевыми структурами генома. Из-за этого появились и ферменты для достройки ССА-участков, и функция добавления неспаренного концевого аденина различными полимеразами. У тех молекул, которые должны были работать рибозимами и не участвовать дальше в копировании, фермент – предшественник РНКазы Р отрезал «клеверный лист». Этот фермент и по сей день имеет в своем составе маленькую РНК-молекулу, что говорит о том, что он очень древний. Отрезанные «клеверные листы» накапливались, и в какой-то момент для них нашлась новая функция: они стали служить адаптерами для прикрепления аминокислот к рибозимам, причем аминокислоты пришивались к ССА-хвосту, а для связывания с рибозимами служили другие участки молекулы, в том числе будущая антикодоновая петля (Szathmá ry, 1993). Итак, вспомогательные молекулы белкового синтеза изначально имели другие функции и могли возникнуть раньше, чем рибосомы. Но как могла появиться сложно устроенная рибосома из десятков взаимосвязанных, подогнанных друг к другу молекул? Допустим, что первая рибосома обходилась без белков (на эту возможность намекает тот факт, что присоединение новой аминокислоты к цепочке до сих пор катализируется РНК). Но рибосомные РНК сами по себе очень сложны. История рибосомных РНК Изучение структуры рибосомной РНК (рРНК) показало, что она могла быть построена постепенно, путем добавления новых блоков к уже существующим. Рибосомная РНК большой субъединицы состоит из шести относительно самостоятельных структурных блоков, или доменов (отмечены римскими цифрами на рис. 13.4). Структура каждого домена определяется прежде всего связями внутри него. В рибосоме домены образуют что-то вроде трехмерного пазла. Причем собрать этот пазл, не ломая детали, можно только в одном порядке, начиная с пятого домена. Это наводит на мысль, что пятый домен древнее всех остальных. Подозрения подтверждаются, если изучить распределение одного из типов связей между участками РНК, так называемых А-минорных контактов. Это музыкальное название обозначает водородные связи между несколькими последовательными аденинами в одноцепочечном участке и гидроксильными группами рибозы в двухцепочечном участке. Такая связь важна для устойчивости одноцепочечного участка, но не влияет на двухцепочечный, поэтому она могла возникнуть, только если одноцепочечная сторона А-минорного контакта возникла позднее, чем двухцепочечная.
На рисунке 13.4 мы видим распределение таких контактов в большой рибосомной РНК кишечной палочки. Видно, что большинство контактов направлены к пятому домену. Это означает, что пятый домен был древнейшим, а остальные части рибосомной РНК надстроились позже (К. Боков, С. Штейнберг, 2009). Некоторые ученые пришли к аналогичным выводам, используя другие признаки: с удалением от пятого домена, в котором находится пептидил-трансферазный центр (именно там и производится присоединение одной аминокислоты к другой в цепочку), уменьшается содержание ионов магния, увеличивается доля белков и повышается упорядоченность их укладки, а в самом пептидил-трансферазном центре белков нет вовсе (Hsiao еt al., 2009). Недавно эти эволюционные реконструкции получили экспериментальное подтверждение: была синтезирована рибосомная РНК длиной 615 нуклеотидов (в 5 раз меньше, чем в обычных бактериальных рибосомах), состоящая из целого пятого домена и древних фрагментов второго и четвертого доменов. Она оказалась способна к взаимодействию с транспортными РНК и к проведению пептидил-трансферазной реакции (Hsiao et al., 2013). Современные рибосомы состоят из двух субъединиц. Если большая непосредственно сшивает аминокислоты, то малая субъединица организует взаимодействие транспортной РНК с матричной, контролирует соответствие кодона антикодону, первой садится на матричную РНК и ищет место начала синтеза белка, а также двигает матричную РНК через рибосому. Молекулы транспортных РНК одним концом (акцепторный стебель) связываются с большой субъединицей, а другим (антикодоновая петля) – с малой. Когда методы определения возраста доменов рибосомной РНК были применены к малой субъединице, результат оказался похож на тот, что мы видели для большой: древнейшей частью молекулы оказался декодирующий центр, в котором происходит связывание антикодоновой петли тРНК с матричной РНК и считывание последовательности матричной РНК. В обоих рибосомных РНК участки, отвечающие за связь субъединиц между собой, моложе, чем пептидил-трансферазный и декодирующий центры (Harish & Caetano-Anolles, 2012). Получается, что две субъединицы рибосомы сначала существовали по отдельности и только потом объединились. Как могла работать древняя большая субъединица рибосомы без белков и без части доменов рРНК? На пятом домене находится пептидил-транферазный центр рибосомы – он присоединяет новые аминокислоты к растущему полипептиду. Однако этот домен не умеет связываться с матричной РНК и потому никак не может выстроить последовательность аминокислот в цепочке белка. Получается, что древняя большая субъединица рибосомы собирала пептиды из аминокислот без участия матричной РНК и использовала какие-то другие механизмы для контроля их последовательности. Дальше в этой главе мы рассмотрим, какие это могли быть пептиды и какими способами рибосома могла контролировать их последовательность до появления матричных РНК. РНК малой субъединицы тоже вела какую-то свою, самостоятельную жизнь до того, как войти в состав рибосомы. Исследователи склоняются к тому, что раньше она работала в качестве РНК-полимеразы или РНК-лигазы, то есть строила новые молекулы РНК из отдельных нуклеотидов или из коротких фрагментов по несколько нуклеотидов. Ее современные функции – движение по матричной РНК и контроль комплементарности кодона с антикодоном – близки к функциям полимераз. Полимеразы тоже движутся по копируемой нити РНК и контролируют комплементарность присоединяемого нуклеотида (Noller, 2010). Так что, похоже, малая субъединица вошла в состав рибосомы, когда происходил переход к белкам, кодируемым при помощи матричных РНК. |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 65; Нарушение авторского права страницы