|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
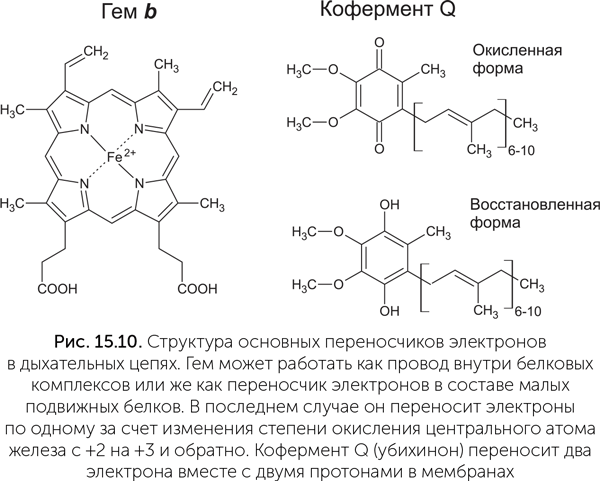
Устройство электрон-транспортных цепей
Процесс, который биохимики называют «электронный транспорт», представляет собой упорядоченное движение электронов от молекул-восстановителей к молекулам-окислителям. Физик назвал бы это проще: «электрический ток». Мембранные электрохимические процессы в клетках отличаются от электрических явлений неживой природы тем, что в клетках переплетаются два разных электрических тока: ионный и электронный. Для ионов вода – проводник, а мембрана – изолятор, ионные токи направлены поперек мембраны через белковые молекулы. Путь электронов сложнее. Привычный нам электрический ток – это движение электронов в металлических проводниках и полупроводниковых кристаллах. В клетках нет металлических проводов, но есть молекулы, выполняющие их функцию внутри больших белковых комплексов. Один из двух типов биологических «проводов» строится из гемов – плоских квадратных молекул с богатой системой двойных связей с атомом железа в центре (рис. 15.10). Гем известен в первую очередь как связывающая кислород «деталь» белка гемоглобина, придающая красный цвет нашей крови. Но в клетках есть множество видов других гем-содержащих белков, цитохромов, которые участвуют в переносе электронов. Цитохромы в составе комплексов электрон-транспортных цепей содержат по несколько гемов, прилегающих друг к другу краями. Цепочка гемов проводит электрический ток за счет подвижных электронов в двойных связях молекул. Ближайшим искусственным аналогом такого проводника является графен (слой графита толщиной в один атом). Второй тип внутрибелковых проводов строится из железосерных кластеров, обычно 4Fe-4S, которые подобны наночастицам полупроводящего минерала пирита (FeS2).
Для переноса электронов через воду нет другого пути, кроме как вместе с ионами. Например, хорошие переносчики получаются на основе металлов переменной валентности – железа и меди. В дыхательной цепи участвует цитохром c – маленький белок с одним гемом, который переносит электрон благодаря окислению и восстановлению атома железа в геме. В электрон-транспортной цепи фотосинтеза есть другой маленький подвижный белок, пластоцианин, который содержит медь. Кроме металлов электроны могут переноситься через воду в комплекте с протонами, образуя атомы водорода. Мы уже много раз упоминали молекулы-переносчики водорода – НАД и ФАД. В мембранах есть аналогичные жирорастворимые переносчики – хиноны. Электроны попадают в дыхательную цепь клеток человека и животных в основном на НАД. Первый комплекс дыхательной цепи, НАДН-дегидрогеназа, принимает атомы водорода от восстановленного НАДН, после чего эти атомы водорода разделяются на части: электроны по железосерным проводам уходят на другую сторону белкового комплекса, а протоны просто выбрасываются в воду. Если для ионного тока в клетке легко указать направление – поперек мембраны, то путь электронного тока описывается сложнее. Можно указать химический состав «плюса» и «минуса» его батарейки (в клетках человека это в основном кислород и НАДН), но оба полюса распределены в объеме клетки. В клеточной электрической цепи на пути от «плюса» к «минусу» чередуются большие неподвижные комплексы дыхательной цепи и маленькие подвижные переносчики. Мы можем указать только порядок молекул, между которыми по цепочке перемещаются электроны. Первый комплекс (НАДН-дегидрогеназа) имеет «разъемы» (сайты связывания) для НАДН и для хинона. Третий комплекс (цитохром bc1) имеет сайты связывания для хинона и подвижного цитохрома с. Наконец, четвертый комплекс (цитохром-с-оксидаза) имеет сайты связывания для цитохрома с и кислорода. Второй номер в этой нумерации достался вспомогательному комплексу, окисляющему янтарную кислоту и отдающему электроны на хинон. В электрической цепи он подключен параллельно первому комплексу. Встречи больших неподвижных комплексов и маленьких подвижных переносчиков происходят благодаря тепловому движению молекул. Каждую секунду каждый комплекс дыхательной цепи сотни раз контактирует с молекулами-переносчиками электронов всех типов, какие только есть в клетке. Представьте себе электрическую цепь, в которой все соединительные провода много раз в секунду выдергиваются из своих разъемов и тыкаются обратно во все разъемы подряд. Примерно так работают электрические цепи в клетках. Чтобы не произошло «короткого замыкания» (переноса электронов с НАДН на кислород напрямую, мимо всех или части комплексов дыхательной цепи), разные комплексы дыхательной цепи должны иметь разные, несовместимые «разъемы» под разные переносчики. Кроме того, эти «разъемы» по-разному расположены относительно мембраны: сайты связывания НАДН находятся на внутренней стороне белковых комплексов, сайты для хинона – в толще мембраны, а для цитохрома с – на внешней стороне комплексов дыхательной цепи. Цитохромные и железосерные провода обычно имеют длину порядка толщины мембраны – 10 нм. Однако бактерии семейства Desulfobulbaceae из тех же деталей строят провода вполне солидных размеров – до 1, 5 см в длину. Эти нитчатые бактерии населяют морское дно и получают энергию, окисляя сероводород кислородом. Цепочки цитохромов на поверхности соседних клеток, соединенных в длинную нить, точно состыкованы друг с другом и проводят электроны по всей длине нити, позволяя бактериям использовать кислород из воды, а сероводород – из глубины осадка (Pfeffer, 2012). Q-цикл Решающее преимущество, благодаря которому протонная энергетика сильно потеснила натриевую в живом мире, – ее универсальность. Она обеспечивается тем, что для протонов (но не для ионов натрия) есть механизм переноса через мембрану, который стыкуется практически с любой окислительно-восстановительной реакцией. Он называется Q-цикл, по главной действующей молекуле – коферменту Q, или убихинону (да, это то самое вещество Q10, которое добавляют в косметику). Молекулы убихинона гидрофобны и находятся в мембранах. Кроме того, хинонное кольцо убихинона может легко присоединять и отдавать два электрона или атома водорода (рис. 15.10). В простейшем варианте Q-цикла перенос протонов происходит за счет того, что один комплекс дыхательной цепи, отдающий электроны на хинон, имеет сайт связывания хинона на внутренней стороне мембраны, а другой, принимающий электроны, – на наружной стороне (рис. 15.11). Хинон может пройти через мембрану, только когда его электрический заряд равен нулю. Иначе говоря, в комплекте с электронами должны быть связаны протоны. Поэтому хинон присоединяет два протона из воды внутри клетки и отдает их в воду снаружи. Ни сам хинон, ни хинон-связывающие сайты ферментов не связаны непосредственно с окислителем и восстановителем, электроны проходят от восстановителя к хинону и от хинона к окислителю по внутрибелковым «проводам» из гемов или железосерных кластеров, никаких движений белковых молекул не требуется. В этой независимости и кроется секрет универсальности убихинонного цикла. Натриевые же насосы вынуждены использовать движения частей белковой молекулы для переноса ионов через мембрану и поэтому труднее перестраиваются на другие реакции.
Подводим итоги: общий предок бактерий и архей имел примитивные мембраны, вероятнее всего, с однохвостыми липидами из терпенового спирта и полярной головки, без глицерола. Эти мембраны пропускали протоны и ионы металлов, но задерживали белки и РНК, поэтому для управляемого выделения белков во внешнюю среду возникли системы секреции. После разделения линий бактерий и архей у них независимо возникли мембраны из липидов с двумя гидрофобными хвостами, непроницаемые для ионов металлов. Выход из исходных геотермальных водоемов в моря с их высокой концентрацией натрия потребовал создания систем откачки натрия из клеток. Так появились натриевые насосы, использующие энергию разных химических реакций. Один из насосов получился из системы секреции белка, в которой застрял секретируемый продукт, превратив ее в роторную Na-движущую АТФазу, и она стала откачивать натрий за счет гидролиза АТФ. Это событие могло произойти два раза независимо в линиях бактерий и архей и дать F– и A/V семейства роторных АТФаз.
Далее горизонтальный перенос генов совместил в одной клетке разные натриевые насосы, в том числе такие, которые создавали больший потенциал, чем роторная АТФаза (рис. 15.12). В этих условиях пригодилось то, что роторная АТФаза обратима: она стала впускать натрий внутрь клетки с синтезом АТФ, и создаваемый химическими насосами натриевый потенциал начал частично расходоваться на синтез АТФ (если взять два разных электрических генератора и соединить их проводами, то тот из них, который давал меньшее напряжение, станет работать электродвигателем). Затем, по мере освоения кислых местообитаний, появились протононепроницаемые мембраны, а мутация Na-АТФазы превратила ее в протонную. Это позволило использовать ее для откачки лишних протонов из клетки за счет энергии АТФ. Другие прокариоты для той же цели откачки лишних протонов приспособили комплексы дыхательной цепи. После этого горизонтальный перенос генов свел в одной клетке протон-движущие комплексы дыхательной цепи и протон-движущую АТФазу. Протонная энергетика благодаря универсальности Q-цикла, позволяющего легко использовать самые разные окислительно-восстановительные реакции, в значительной степени вытеснила натриевую.
Глава 16 |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 65; Нарушение авторского права страницы