|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Другие возможные источники генов эукариот
Мы определили два основных источника генов эукариот: это альфа-протеобактерия, давшая начало митохондриям, и архея, ставшая ее хозяином в симбиозе. Однако этих источников недостаточно: в геномах эукариот есть много генов, которые похожи на гены разных бактерий, но отсутствуют у альфа-протеобактерий. Кроме митохондрий у эукариот есть другая органелла, происходящая от симбионта, – хлоропласт водорослей и растений. Хлоропласты происходят от цианобактерий и, по некоторым гипотезам, могли быть у общего предка всех эукариот и утратились во всех ветвях, кроме водорослей и происходящих от них растений. Может быть, они внесли свой вклад в геном всех эукариот? Сравнение геномов показывает, что это не так. У эукариот насчитывается более 200 генных семейств бактериального происхождения, которых нет ни у альфа-протеобактерий, ни у цианобактерий, и, следовательно, приобретенных другими путями. Функции этих генных семейств чаще всего связаны с регуляцией и передачей сигналов, тогда как гены альфа-протеобактериального происхождения связаны прежде всего с обменом веществ (рис. 18.5). Вклад цианобактерий заметен только в ядерных геномах водорослей и растений. Некоторые из белков эукариот, хотя имеют альфа-протеобактериальных родственников, но ближе к белкам других групп бактерий. Например, все ферменты гликолиза эукариот наиболее сходны с белками бактерий-бродильщиков рода Clostridium.
Несколько белковых семейств эукариот, связанных со слиянием и отшнуровкой мембранных пузырьков, имеют архейное происхождение – например, Adaptin N, Syntaxin, AdAR repeat. К сожалению, их функции в клетках архей неизвестны. Помимо самих митохондрий, организующих кислородное дыхание, для приспособления к кислородной среде нужны белки, защищающие цитоплазму от повреждений, такие как оксигеназы, пероксидазы, каталаза. Эти белки эукариот получены от бактерий, причем не родственных митохондриям. Чего и следовало ожидать – ведь, чтобы получать выгоду от аэробного дыхания, надо сначала научиться защищаться от токсического действия кислорода, которое проявляется уже при очень малых его концентрациях. Мембраны эукариот, подобно бактериальным, состоят из сложных эфиров жирных кислот. Ферменты биосинтеза липидов, соответственно, могли быть получены только от бактерий. По своим аминокислотным последовательностям они ближе всего к ферментам альфа-протеобактерий, а значит, получены от предков митохондрий. Другая важная группа липидов, инозитол-фосфаты, унаследована эукариотами от архей. Инозитол-фосфатные липиды составляют небольшую долю мембран эукариот, но важны как метки мембран разного назначения, регуляторы состояния мембранных белков, слияния и разделения мембранных пузырьков и даже регуляторы работы генов (на стадии созревания матричных РНК и экспорта их из ядра). Важно, что древние и универсальные мембранные белки, такие как роторная АТФаза или сигнал-распознающая частица, встраивающая новые белки в мембрану, у эукариот похожи на архейные, но работают в мембране, состоящей в основном из липидов бактериального типа. Значит, в эволюции эукариот был этап замены мембранных липидов с архейных на бактериальные. Японские биохимики воспроизвели промежуточную стадию этого процесса, создав трансгенную кишечную палочку с генами синтеза архейных липидов (Shimada, Yamagishi, 2011). Оказалось, что смесь липидов разного происхождения практически не влияет на жизнеспособность клеток при любых соотношениях бактериальных и архейных липидов. Отличительной чертой мембран эукариотических клеток являются вспомогательные липиды – стеролы, такие как холестерол. Стеролы повышают текучесть мембран и поэтому важны для фагоцитоза и транспорта мембранных пузырьков. Выходит, что стеролы, вероятно, появились раньше, чем митохондрии. Предшественником всех стеролов является терпеновый углеводород сквален, который мы уже упоминали в главе 15 как добавку к липидам у бактерий, живущих в щелочных средах. В качестве промежуточного продукта обмена веществ сквален распространен гораздо шире, он синтезируется у многих бактерий и архей. Однако дальнейшие стадии синтеза стеролов характерны только для эукариот. На первом шаге сквален-монооксигеназа катализирует присоединение кислорода к концевой двойной связи сквалена, образуя сквален-2, 3-эпоксид. Для этой реакции требуется молекулярный кислород, хотя и в очень малой концентрации. Затем ланостерол-синтаза катализирует восстановление эпоксида, при этом неустойчивый продукт восстановления – радикал сквалена – перестраивает двойные связи в замкнутые кольца, образуя ланостерол (рис. 18.6). Ланостерол дает начало всем другим стероидам путем модификации боковых групп.
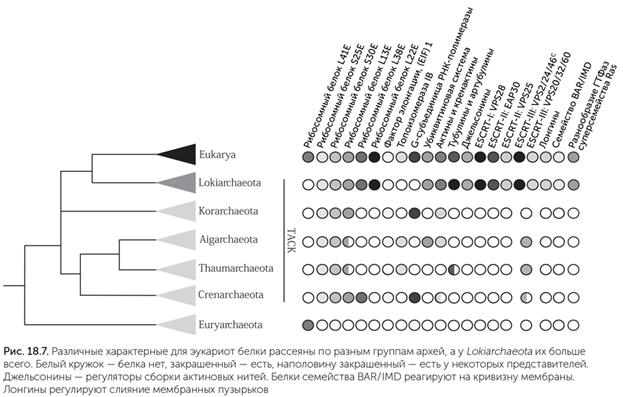
Из-за крайней важности стеролов для эукариотической клетки происхождение ферментов их биосинтеза внимательно исследовалось. Сходные ферменты, синтезирующие те же стеролы, были обнаружены у некоторых гамма-протеобактерий, например Methylococcus capsulatus (Lamb et al., 2007), у Gemmata obscuriglobus из планктомицетов (Pearson et al., 2003) и у ряда микобактерий, например Plesiocystis pacifica и Stigmatella aurantiaca. Однако подробное исследование показало, что все эти ферменты бактерий получены горизонтальным переносом от эукариот уже после расхождения линий растений и животных (Desmond, Gribaldo, 2009). Первый шаг биосинтеза стеролов – присоединение кислорода к сквалену – мог быть поначалу одним из способов защиты от молекулярного кислорода, и лишь потом нашлось применение полученному сквален-2, 3-эпоксиду. Большая часть эукариотических генов бактериального происхождения, не принадлежавших предкам митохондрий и пластид, кодирует защитные, рецепторные, транспортные, сигнальные и регуляторные белки. Сюда относятся, например, белки теплового шока Hsp90, белки устойчивости к тяжелым металлам TerC и Ttg2 и др. Что интересно, белки, которые у эукариот обеспечивают взаимодействие между компонентами клетки, у бактерий нужны для связей между разными клеткам в сообществе. Например, Tim44 эукариот необходим для транспорта белков через внутреннюю мембрану митохондрий из цитоплазмы, а его бактериальные родственники экспортируют белки из клеток во внешнюю среду. Это свидетельствует о том, что при возникновении эукариотической клетки под одной мембраной оказались уже отработанные системы связи клеток в сообществе. В целом можно сказать, что те белки эукариот, которые получены от других бактерий (не предков митохондрий и пластид), не имеют какого-то одного источника. Скорее, они были получены от множества разных бактерий путем горизонтального переноса генов. Гипотезы, в которых до симбиоза с протеобактериями было слияние клеток какой-то другой бактерии с археей, не поддерживаются этими геномными данными. Архейный предок эукариот Хотя участие какой-то археи в происхождении эукариот давно стало очевидно, выделить конкретную группу архей, к которой мог относиться предок эукариот, оказалось не так просто. Многие компоненты эукариотной клетки были найдены у отдельных видов архей, но это были разные виды, относящиеся к разным крупным группам архей. Так обстоит дело, например, с цитоскелетными белками тубулинами. Давно известны белки FtsZ, отдаленно похожие по последовательности на тубулин и широко распространенные у бактерий и архей. Они собираются в пластины и трубки и необходимы для деления прокариотических клеток. В 2012 году были найдены артубулины – близкие родственники тубулина – у двух видов архей рода Nitrosoarchaeum, принадлежащих к группе Thaumarchaeota. Их функция пока неизвестна (Yutin, Koonin, 2012). Аналогично у бактерий и архей широко распространены белки, отдаленно сходные с актином; их называют MreB. Они собираются в тонкие нити и участвуют в перетяжке делящейся бактериальной клетки. Другие дальние белковые родственники актина кодируются плазмидами (маленькими независимыми молекулами ДНК у бактерий) и служат для их расхождения при делении клетки. В 2009 году у нескольких видов группы Crenarchaeota были обнаружены кренактины, весьма похожие на актин эукариот (Ettema et al., 2011). Кренактины образуют нити, которые придают клеткам палочковидную форму, часто с разветвлениями. По своей последовательности кренактины близки не только к актинам, но и к белкам Arp2/Arp3 (Actin-related proteins), которые у эукариот образуют разветвления актиновых нитей. В некоторых случаях совместная полимеризация актина и Arp2/Arp3 на внутренней стороне клеточной мембраны достаточна для отшнуровки фагоцитозного пузырька (Yutin et al., 2009). Наряду с артубулинами у Thaumarchaeota обнаружились гены еще одной эукариотической системы – ESCRT-III (endosomal sorting complex required for transport). Эта группа белков связана с мембранами комплекса Гольджи, эндоплазматического ретикулума и вакуолей; она участвует в сортировке содержимого мембранных пузырьков. В клетках Thaumarchaeota белки этого комплекса необходимы для деления клеток, работая вместо отсутствующего у них FtsZ (Makarova et al., 2010). В геноме Caldiarchaeum subterranum, прочитанном в 2010 году, были обнаружены гены важной для эукариот системы убиквитиновой модификации белков. Эта система пришивает убиквитин – маленький белок – к различным клеточным белкам. Убиквитин служит обычно меткой для уничтожения белков. Кроме собственно убиквитина (Ub) в нее в минимальном варианте входит три фермента – Е1, Е2 и Е3, катализирующие разные стадии присоединения Ub. У бактерий известны отдаленные родственники Е1 и Е2, но их функции не связаны с убиквитином, они катализируют включение серы при синтезе витамина В1 и молибденовых кластеров ферментов. Меченые убиквитином белки обычно разрушаются в протеасомах – специальных белковых комплексах для выборочного уничтожения дефектных и ненужных белков. Функциональное ядро протеасомы состоит из 28 белковых молекул, собранных в четыре кольца из семи субъединиц; кроме того, в ее работе принимают участие дополнительные регуляторные субъединицы. Протеасомы были известны ранее у некоторых бактерий (Mycobacterium tuberculosum) и архей (Haloferax volcanii), но убиквитиновой системы у этих видов нет. Протеасомы есть и у Caldiarchaeum subterranum, они похожи на эукариотические также по наличию белка RPN11, который опознает убиквитиновые метки на белках (Nunoura et al., 2011). Почти все, что мы знаем об устройстве и образе жизни бактерий и архей, ученые узнали, выращивая и изучая микробов в лаборатории. Те микробы, для которых не удалось подобрать условия выращивания, долго оставались неизвестными. В XXI веке появились методы метагеномики – анализа всей ДНК, содержащейся в природной пробе (вода, почва, ил). Метагеномика позволяет оценить разнообразие микробов в пробе без культивирования. Используя метагеномный способ, ученые быстро выяснили, что микробное разнообразие в природе огромно. Более 99 % видов микробов, которые есть в природе, никогда не вырастали в лаборатории, и до появления метагеномики об их существовании можно было только догадываться. Хуже того, половина крупных групп бактерий и архей не имеет ни одного культивируемого представителя и известна только по метагеномным данным. Ученые подозревали, что среди них скрываются и близкие родственники эукариот. И вот в 2015 году такие археи, близкие к эукариотам, были найдены (Spang et al., 2015). Норвежская экспедиция, изучавшая геотермальное поле Замок Локи (Loki's castle) в Северной Атлантике, собрала пробы донных осадков, обильно заселенных бактериями и археями. Анализ ДНК в этих пробах показал, что в сообществе преобладает один вид архей, относящийся к некультивируемой группе архей DSAG (deep-sea Archaea group). На его долю приходилось более 10 % клеток, а значит, появилась возможность прочитать не только гены рибосомных РНК, по которым устанавливают родственные связи микробов, но и собрать весь его геном. Этот вид, который получил временное название Lokiarchaeum, оказался ближе к эукариотам, чем все известные ранее археи. Его геном имеет размер более 5 млн пар нуклеотидов и кодирует 5381 белок, поэтому является одним из самых сложных геномов архей. Среди этих белков есть пять актиноподобных белков, более похожих на актины и ARP эукариот, чем кренактины (рис. 18.7). Также Lokiarchaeum обладает большим набором сигнальных белков суперсемейства Ras – 92 разновидностями! Эти белки в клетках эукариот регулируют перестройки цитоскелета, транспорт пузырьков, передачу сигналов между мембраной, цитоплазмой и ядром, деление клеток и многие другие функции. Белки Ras человека очень хорошо изучены, потому что мутации в них часто приводят к неограниченному делению клеток и раку. Хотя отдельные Ras-подобные белки встречаются в разных группах бактерий и архей, только Lokiarchaeum имеет их большой набор. При этом часть его Ras-белков попадает в отдельные эукариотные подсемейства этого суперсемейства. Еще у Lokiarchaeum есть большой набор генов системы ESCRT-III; в отличие от Thaumarchaeota у него уже произошло выделение нескольких эукариотных подсемейств этих генов. Также у него появляются гены комплексов ESCRT-I и ESCRT-II, которые у эукариот взаимодействуют с ESCRT-III, собирая вместе и загружая в мембранные пузырьки меченные убиквитином белки, подлежащие разборке на аминокислоты.
По набору генов, связанных с цитоскелетом и управлением мембранами, похоже, что Lokiarchaeum и его общий с эукариотами предок могут иметь способность к фагоцитозу. К сожалению, Lokiarchaeum не растет в лаборатории, поэтому пока нет возможности проверить, есть ли у него фагоцитоз на самом деле. Зато в тех же пробах из Замка Локи есть в меньшем количестве ДНК других локиархеот, и одна из них, Loki3, еще ближе к эукариотам. Через пару лет и ее геном, скорее всего, будет прочитан. |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 56; Нарушение авторского права страницы