|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Бактериофаги: основные морфологические формы, структура фагов. Вирулентные и умеренные фаги. Этапы взаимодействия фага с клеткой.
Структура типичного миовируса бактериофага (рис. 1). Бактериофаги представляют собой наиболее многочисленную, широко распространенную в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц. В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения. Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем». Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 1024 бактерий. Это означает, что постоянный перенос генетического материала распределяется между бактериями, обитающими в сходных условиях. Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды. Бактериофаги различаются по химической структуре, типу нуклеиновой кислоты[2], морфологии и характеру взаимодействия с бактериями. По размеру бактериальные вирусы в сотни и тысячи раз меньше микробных клеток.
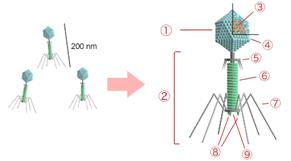
Рис. 2. Строение бактериофага 1 – головка, 2 – хвост, 3 – нуклеиновая кислота, 4 – капсид, 5 – «воротничок», 6 – белковый чехол хвоста, 7 – фибрилла хвоста, 8 – шипы, 9 – базальная пластинка. Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2 – 4 раза больше диаметра головки. В головке содержится генетический материал – одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окруженная белковой или липопротеиновой оболочкой – капсидом, сохраняющим геном вне клетки. Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Обычно углы состоят из пентамеров белка, а опора каждой стороны из гексамеров того же или сходного белка. Более того, фаги по форме могут быть сферические, лимоновидные или плеоморфные. Хвост представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Существуют также бактериофаги с коротким отростком, не имеющие отростка и нитевидные. Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они переносят всю информацию для запуска собственной репродукции в соответствующем хозяине, у них отсутствуют механизмы для выработки энергии и рибосомы для синтеза белка. У некоторых фагов в геноме содержится несколько тысяч оснований, тогда как фаг G, самый крупный из секвенированных фагов, содержит 480 000 пар оснований — вдвое больше среднего значения для бактерий, хотя всё же недостаточного количества генов для важнейшего бактериального органоида как рибосомы. По характеру взаимодействия с клеткой бактерии бактериофаги делятся на вирулентные и умеренные. Вирулентные фаги всегда лизируют клетку бактерии. Умеренные фаги могут вызвать лизис клетки бактерии, но могут перейти и в неинфекционную форму. В этом случае молекула ДНК фага прикрепляется к ДНК бактерии и передается с нею дочерним клеткам. Фаг, существующий в такой форме, называется профагом. Сравнительно недавно стало известно, что включение вирусной ДНК в бактериальную происходит путем кроссинговера между хромосомами бактерии и вируса. Хромосома вируса принимает кольцевую форму и прикрепляется к определенному локусу хромосомы бактерии. Затем хромосомы бактерии и вируса разрываются, концы их соединяются крест-накрест и профаг оказывается включенным в хромосому клетки хозяина. В этом случае профаг является как бы частью ДНК бактерии и вместе с ней реплицируется. Клетки бактерии, имеющие в своей хромосоме профаг, называются лизо-генными, а явление совместного существования ДНК бактерии и профага называется лизогенией.
58. Лизогения λ -типа и P-типа. Регуляция транскрипции генома бактериофага λ. Лизогения (от греч. lý sis — разложение, распад и...geneia — происхождение, создание) генетически обусловленная способность бактерий лизироваться с выделением бактериофага (См. Бактериофаги) через ряд поколений после непосредственного заражения им. Теория Л. разработана в 1950 французскими учёными А. Львовым и А. Гутман, показавшими, что лизогенное состояние связано с присутствием в клетках бактерий потенциально инфекционной структуры — Профага. В каждом поколении лизогенных бактерий подвергается Лизису очень небольшая часть клеток (Лизогения1 клетка на миллион) с освобождением от 70 до 150 частиц так называемого умеренного фага. Частота перехода профага в инфекционное состояние (индукция профага) может быть увеличена рядом агентов (например, ультрафиолетовыми лучами). После заражения бактериальной клетки умеренным фагом процесс инфекции может развиваться по одному из двух направлений: по пути литического цикла, который так же, как и при заражении бактерий вирулентными фагами, заканчивается лизисом клеток и выходом потомства фага в окружающую среду; по пути лизогенизации, когда в результате биосинтетических процессов в клетке вырабатывается иммунитет к инфицирующему фагу, фаговая ДНК включается в ДНК бактерии и в дальнейшем реплицируется вместе с ней как её составная часть (профаг), а бактерия выживает и становится лизогенной. Судьба клетки решается на первых этапах инфекции и зависит главным образом от времени формирования иммунитета. Если состояние иммунитета наступает раньше, чем развитие инфекции достигнет стадии, необратимо ведущей к лизису, то может осуществиться лизогенизация. В Геноме бактерий могут содержаться одновременно профаги нескольких разных фагов (пол и лизогения). В этом случае клетка обладает иммунитетом в отношении всех этих фагов. В результате лизогенизации может произойти изменение некоторых свойств бактериальной клетки (т. н. Лизогенная конверсия), обусловленное приобретением бактерией новой генетической информации. Искусственно полученные лизогенные бактерии по своим свойствам не отличаются от лизогенных бактерий, найденных в естественных условиях. У небольшой части потомства лизогенной клетки происходит «исцеление» — потеря профага. Утратившие профаг клетки дают начало нелизогенным линиям. Частота этого процесса может быть увеличена, например, действием ультрафиолетовых лучей. Т. о., Л. — сложная форма вирусной инфекции у бактерий, при которой от момента заражения бактерий фагом до лизиса клетки проходит большое число клеточных поколений. В вирусных и клеточных системах молекулярные механизмы транскрипции принципиально сходны. Отличие заключается в существовании различных способов регуляции транскрипции вирусных геномов. Необходимость такой регуляции определяется разной потребностью в вирусоспецифических белках. Структурные белки, как правило, требуются в больших количествах, чем белки-ферменты. Кроме того, на ранних стадиях инфекции нужны белки, обеспечивающие репликацию вирусного генома, а на поздних — структурные белки. Поэтому целесообразно, чтобы разные вирусные гены считывались с разной эффективностью, и эта эффективность менялась во времени. Процесс транскрипции регулируется на уровне транскриптона (оперона) за счет работы репрессоров и активаторов белковой природы и энхансеров (усилителей), которые представляют собой определенные короткие последовательности геномной нуклеиновой кислоты. Транскрипция регулируется количественно и качественно и осуществляется как клеточными, так и вирус-специфическими механизмами. У вирусов установлено существование целого ряда способов регуляции транскрипции. Временной тип регуляции. У ДНК-содержащих вирусов существует три периода транскрипции: сверхранний, ранний и поздний. При сверхранней и ранней транскрипции считываются сверхранние и ранние гены, при поздней — поздние гены. Количество транскриптов поздних генов превышает количество ранних. Многие сверхранние мРНК являются генами NS белков-ферментов и регуляторов транскрипции и репликации. Поздние мРНК являются генами структурных белков. Фактором регуляции транскрипции у ядерных вирусов является транспорт мРНК в цитоплазму. Каскадный тип регуляции транскрипции генов. Суть такой регуляции заключается в том, что продукты сверхранней транскрипции, например α -белки, необходимы для транскрипции другой группы генов, кодирующих β -белки, которые, в свою очередь, включают транскрипцию следующей группы генов — γ -белков. Полярный тип регуляции определяется порядком расположения генов в геноме. Количество синтезируемых молекул полипептида зависит от расстояния между геном и промотором. Вдоль генома (-)РНК вирусов существует как бы градиент эффективности транскрипции. Чаще транскрибируются гены 3'-региона, реже — гены 5'-конца. Взаимное расположение и сила регуляторных сигналов. Считывание или несчитывание транскрибируемого участка матрицы зависит от свойств и расположения регуляторных сигналов — промоторов (обеспечивают начало транскрипции) и терминаторов (обеспечивают прекращение транскрипции). Основа регуляции — взаимное расположение регуляторных сигналов и их сила. Активность сигналов может меняться во времени. Характер образования транскриптов и способ регуляции зависят от того, имеем ли мы дело с вирусами прокариот или эукариот. Напомним, что в клетках прокариот возможна множественная инициация трансляции на полицистронной матрице, тогда как в клетках эукариот на РНК реализуется только одна точка инициации трансляции и эта мРНК функционально моноцистронна. Ограничения, накладываемые клеткой хозяина, в первую очередь сказываются на механизмах транскрипции и посттранскрипционного созревания мРНК. Приведем конкретные примеры способов регуляции транскрипции вирусных геномов в клетках прокариот и эукариот. Самый простой способ регуляции транскрипции в клетках прокариот установлен у фагов 1М13 и fd, где разная степень экспрессии фаговых генов регулируется за счет расположения и силы промоторов. За счет наличия «сильных» промоторов активно транскрибируются гены, кодирующие основной структурный белок капсида и ДНК-связывающий белок. В то же время, геномная последовательность, кодирующая минорные вирусные белки, имеет промоторы, отнесенные к разряду «слабых». Более сложная регуляция транскрипции генов наблюдается у фага λ, имеющего, как минимум, три типа регуляции транскрипции: 1) ретро-регуляция — осуществляется при участии нуклеотидных последовательностей, расположенных за транскрибируемым геном. Этот участок комплементарен предшествующему участку гена и в образовавшемся транскрипте возникает внутримолекулярная двухнитевая структура, которая впоследствии разрушается РНКазой III; 2) аутогенная регуляция — регуляция активности гена при помощи продукта этого же гена; 3) индукция профага — наблюдается в результате инактивации репрессора. Для бактериофагов показана реализация временного типа регуляции транскрипции, что связано с существованием ранних, средних и поздних генов и соответствующих им промоторов. Так, у фага Т4 структура ранних промоторов близка к таковой промоторов клетки хозяина и именно они сразу узнаются клеточной РНК-полимеразой. Последующая активация средних генов связана с фагоспецифическим белком — продуктом трансляции раннего гена. Система регуляции транскрипции генов фага Т4 включает еще один уникальный механизм — ковалентную и нековалентную модификацию РНК-полимеразы, способствующую узнаванию ею поздних промоторов. Модифицированная РНК-полимераза перестает узнавать промоторы ранних генов. Еще один способ временной регуляции наблюдается у фага Т7. Суть этого способа заключается в том, что одним из продуктов ранних генов, транскрибированных клеточной РНК-полимеразой, является фаговая РНК-полимераза, которая узнает уже другой набор промоторов и транскрибирует поздние гены. Регуляция транскрипции вирусных геномов в эукариотических клетках осуществляется с помощью более сложных механизмов. Кроме промоторов и терминаторов транскрипционная система дополняется новыми регуляторными элементами — энхансерами (усилители), а также и разнообразными способами процессинга первичных транскриптов. В данном разделе мы не станем останавливаться на конкретных способах регуляции транскрипции генов вирусов эукариот, которые, в общих чертах, сходны с перечисленными выше. Как дополнение, рассмотрим процессинг первичных транскриптов на примере ядерного вируса эукариот — аденовируса. Процессинг — это посттранскрипционные изменения первичных транскриптов или созревание мРНК, включающее кэпирование 5'-конца, полиаденилирование 3'-конца и сплайсинг. У аденовируса лишь кэпирование идет эффективно на разных стадиях репродукции и происходит до завершения синтеза транскрипта. Большой вклад в регуляцию экспрессии аденовирусного генома вносит альтернативное полиаденилирование. Особенно наглядно это видно при образовании поздних мРНК. В первичном транскрипте поздней области генов есть 5 участков, несущих сигнал полиаденилирования (гексануклеотид AAUAAA). Полиаденилирование может произойти в любом участке и из первичного транскрипта может образоваться только одна из 5-ти возможных классов мРНК. От выбора того или иного участка полиаденилирования зависит относительная концентрация той или иной мРНК. Подавляющее большинство кэпированных и полиаденилированных транскриптов аденовирусного генома подвергается альтернативному сплайсингу – удалению различных участков первичного транскрипта, что осуществляется при помощи клеточных механизмов. Наличие альтернативного сплайсинга и альтернативного полиаденилирования при процессинге первичных транскриптов вирусов эукариот определяется моноцистронностью эукариотических мРНК.
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 93; Нарушение авторского права страницы
 Бактериофаги (фаги) (от др.-греч. φ ᾰ γ ω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис[1]. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.
Бактериофаги (фаги) (от др.-греч. φ ᾰ γ ω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис[1]. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.