|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ИСТОРИЯ РАЗВИТИЯ УЧЕНИЯ О БИООКИСЛЕНИИ.
А. Лавуазье в конце XVIII века показал, что животный организм потребляет из воздуха кислород и выделяет углекислый газ. Сделал вывод, что горение и окисление - это одно и то же, что биологическое окисление представляет собой " медленное горение", происходящее в присутствии воды и при низкой температуре. В конце XIX века русские исследователи А.Н. Бах и В.И.Палладин, работая независимо друг от друга, предложили 2 основные теории для объяснения процессов, протекающих в ходе биологического окисления. 1-я теория: А.Н.Бах (1857-1946) полагал, что в живых клетках существуют особые ферменты - " оксигеназы", которые взаимодействуют с кислородом, образуя перекиси. Сам кислород является не очень активным окислителем. Зато перекиси (" активный кислород" ) являются очень сильными окислителями и способны передавать кислород окисляемому веществу. Эта теория известна как " перекисная" или " теория активации кислорода".
2-я теория: В.И. Палладин (1859-1922) создал теорию " активации водорода". Считал, что универсальным путем окисления является отнятие от веществ (субстратов) водорода с участием специальных ферментов - хромогенов. После этого водород, по Палладину, может передаваться или на молекулу кислорода с образованием воды, или на другие молекулы, восстанавливая их.
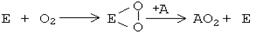
Впоследствии теория В.И.Палладина блестяще подтвердилась для процессов митохондриального окисления, а ферменты, принимающие непосредственное участие в отнятии водорода от субстратов, в настоящее время называются дегидрогеназами. СОВРЕМЕННАЯ ТЕОРИЯ БИООКИСЛЕНИЯ Согласно СОВРЕМЕННОЙ ТЕОРИИ БИООКИСЛЕНИЯ в нашем организме окисление может происходить двумя способами: 1. Путем отнятия водорода от окисляемого субстрата: сюда относятся МИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ и ВНЕМИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ ОКСИДАЗНОГО ТИПА. 2. Путем присоединения кислорода к окисляемому субстрату - так происходит внемитохондриальное ОКИСЛЕНИЕ ОКСИГЕНАЗНОГО ТИПА (старое название - МИКРОСОМАЛЬНОЕ окисление). МИТОХОНДРИАЛЬНОЕ ОКИСЛЕНИЕ (МтО). Система митохондриального окисления - мультиферментная система, постепенно транспортирующая протоны и электроны на кислород с образованием молекулы воды. Все ферменты митохондриального окисления встроены во внутреннюю мембрану митохондрий. Только первый переносчик протонов и электронов - никотинамидная дегидрогеназа расположена в матриксе митохондрии. Этот фермент отнимает водород от субстрата и передает его следующему переносчику. Полный комплекс таких ферментов образует " дыхательный ансамбль" («дыхательную цепь»), в пределах которого атомы водорода отнимаются от субстрата, затем передаются последовательно от одного переносчика к другому, и, наконец, передаются на кислород воздуха с образованием воды. Существует строгая последовательность работы каждого звена в цепочке переносчиков. Эта последовательность определяется величиной РЕДОКС-ПОТЕНЦИАЛА (ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНОГО ПОТЕНЦИАЛА, сокращенно - ОВП ) каждого звена. ОВП - это химическая характеристика способности вещества принимать и удерживать электроны. Выражается в вольтах (V). Вещества с положительным ОВП окисляют водород (отнимают от него электроны), вещества с отрицательным ОВП окисляются самим водородом. Самый низкий ОВП имеет начальное звено цепи, самый высокий - у кислорода, расположенного в конце цепочки переносчиков. Таким образом, передача водорода идет от более низкого к более высокому ОВП. Перенос водорода и электронов возможен только в одном направлении - в порядке возрастания их ОВП: от -0.32V у никотинамидных дегидрогеназ (первого компонента главной цепи МтО) до 0.82V у О2, обладающего самым высоким редокс-потенциалом. На одной из стадий происходит разделение атомов водорода на Н+ и электроны. Протоны остаются временно в окружающей среде, а электроны идут дальше по цепи и в ее конце используются для активации О2. Кислород является конечным акцептором электронов. O2 + 4e -----> 2O-2 (полное восстановление кислорода) Все реакции, происходящие в дыхательной цепи, сопряжены. Переносчики водорода и электронов расположены в строгом порядке, в соответствии с величиной их редокс-потенциала. В настоящее время различают три варианта дыхательных цепей: 1) ГЛАВНАЯ (ПОЛНАЯ) ЦЕПЬ 2) УКОРОЧЕННАЯ (СОКРАЩЕННАЯ) ЦЕПЬ 3) МАКСИМАЛЬНО УКОРОЧЕННАЯ (МАКСИМАЛЬНО СОКРАЩЕННАЯ) ЦЕПЬ.
Сначала разберем их строение на примере главной дыхательной цепи. ГЛАВНАЯ ДЫХАТЕЛЬНАЯ ЦЕПЬ Главная дыхательная цепь - это три мультиферментных комплекса, встроенных во внутреннюю мембрану митохондрии. Обозначаются они латинскими цифрами – I, III и IV. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 492; Нарушение авторского права страницы