|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Глава 6. Адаптивная морфология организмов
Среди приспособлений животных и растений к среде немаловажную роль играют морфологические адаптации, т. е. такие особенности внешнего строения, которые способствуют выживанию и успешной жизнедеятельности организмов в обычных для них условиях. «Целесообразность» строения живых существ долго не могла получить материалистического объяснения. Ч. Дарвин, выдвигая теорию естественного отбора как главного фактора эволюции, обратил внимание на то, что результатом естественного отбора может быть не только нарастание различий между близкими видами, но и выработка у неродственных форм внешнего сходства, если эти виды ведут сходный образ жизни в близких условиях среды. Этот процесс получил название конвергенции. Конвергенция признаков у разных форм в наибольшей мере затрагивает те органы, которые находятся в непосредственном соприкосновении с внешней средой (рис. 67). Внутренние черты строения организмов, их общий план строения остаются при этом неизменными, отражая родство и происхождение видов. Формообразующая роль факторов среды, т. е. влияние их на морфологию организмов, наглядно выступает при изучении роли влажности, температуры, движения воды и воздуха, плотности среды, объема пригодного для жизни пространства и др. Чем жестче физические условия среды, тем ограниченнее пути приспособления к ней. Одинаковые принципы освоения среды ведут к выработке сходных морфологических адаптаций у разных видов, даже сильно различающихся по плану своего строения. На форму быстро движущегося в жидкости тела накладывают жесткие ограничения законы физики. На него действует лобовое сопротивление, которое зависит от природы жидкости, скорости движения, площади проекции тела перпендикулярно направлению движения и длины тела в направлении потока. Для жидкости с такой вязкостью и плотностью, как вода, лобовое сопротивление для тел одинакового объема минимально, если отношение длины к наибольшему диаметру равно примерно 4, 5. Хорошие пловцы среди животных обладают именно такими пропорциями, хотя достигают этого разными средствами. Плывущие кальмары так складывают щупальца, что тело становится торпедообразным, с оптимальным соотношением длины и диаметра, их скорость достигает 41 км/ч. У рыб большое значение имеет форма хвостового плавника; тюлени соединяют и вытягивают задние ласты. По пропорциям тела можно судить о максимально возможной скорости животного в воде (рис. 68). Дельфин-белобочка может плыть со скоростью 44 км/ч, тогда как тюлени, пропорции которых менее оптимальны, – не более 20 км/ч. Обтекаемость тела, таким образом, – необходимый путь приспособления к быстрому движению в воде.
Рис. 67. Конвергентное развитие поверхностей у животных, способных к планирующему полету
Рис. 68. Различные формы тела рыб (по Г. В. Никольскому, 1974): 1– стреловидная (сарган); 2 – торпедовидная (скумбрия); 3 – сплющенная с боков (лещ); 4 – тип луны-рыбы; 5 – тип камбалы, 6 – змеевидная (угорь); 7 – лентовидная (сельдяной король); 8 – плоская (скат); 9 – шаровидная (кузовок)
Среди самых разнообразных по внешнему облику планктонных организмов (одноклеточных водорослей, кишечнополостных, ракообразных, моллюсков, червей, личинок разных групп беспозвоночных, пелагической икры рыб и т. д. (см. рис. 39) мы встречаем три описанных выше способа приспособления к парению: 1) уменьшение объема тела; 2) развитие разнообразных выростов; 3) увеличение содержания в теле воды, жиров и газообразных продуктов. Во всех случаях достигается один эффект: уменьшение отношения массы тела к его площади, что обеспечивает уравновешивание сил тяжести и трения о воду. Форма планктеров и здесь играет большую роль в обеспечении их образа жизни, но отвечает другим (по отношению к нектерам) требованиям законов физики – необходимости увеличения относительной поверхности тела. Это может достигаться разными путями: уменьшением размеров организмов, уплощением, удлинением, умножением числа различных выступов, щетинок и т. п. Таким образом, общее число возможных морфологических приспособлений к одной и той же среде диктуется как ее свойствами, так и способами ее освоения. Морфологический тип приспособления животного или растения к основным факторам местообитания и определенному образу жизни называют жизненной формой организма.
Жизненные формы растений Понятие жизненной формы зародилось при изучении растительного покрова. Сам термин вошел в науку только в конце XIX в., но большое разнообразие форм растений в природе, определяющих ландшафты разных территорий, привлекало ботаников еще в древности. Древнегреческий ученый и философ, ученик и друг Аристотеля, Теофраст (Феофраст) более чем за три столетия до нашей эры в произведении «Исследования о растениях» систематизировал накопленные знания по морфологии растений, выделил деревья, кустарники, полукустарники, травы и описал их. Деревья он характеризовал как растения со стволом, кустарники – со множеством веток, отходящих прямо от корня, полукустарники – как растения, которые дают от корня много стеблей и множество веточек. Травы он объединял в группы по длительности жизни, характеру побегов, листьев, корневых систем, наличию луковиц и клубней. Он отмечал зависимость формы роста от климата, почвы, способов возделывания. Группы жизненных форм служили Теофрасту, как и многим ботаникам нового времени, основой для систематики растений. Основоположник ботанической географии немецкий ученый А. Гумбольдт (1769–1859) в результате знакомства с растительностью разных континентов в начале XIX в. выделил сначала 16, а затем 19 «основных форм», различающихся физиономически: пальмы, бананы, мальвовые и баобабовые, вересковые, кактусовые, орхидеи, казуариновые, ароидные, лианы, алоэ, злаки, папоротники, лилейные, ивовые, миртовые, меластомовые, хвойные, мимозовые, лотосовые. Это не систематические группы. В основе выделения «основных форм» лежит сходство во внешнем облике. Форма злаков, например, включает осоки и представителей других семейств с узкими листьями. В форму алоэ включены агавы, ананас, панданусовые и др. Сходство внешнего облика разных растений Гумбольдт объяснял влиянием климатических, почвенных условий, высотой над уровнем моря и проч. Термин «жизненная форма» предложен в 1884 г. датским ботаником Е. Вармингом. Под жизненной формой Варминг понимал «форму, в которой вегетативное тело растения (индивида) находится в гармонии с внешней средой в течение всей его жизни, от колыбели до гроба, от семени до отмирания». Из определения следует, что жизненная форма отражает приспособленность растения ко всему комплексу факторов внешней среды во все периоды его жизни. Датский ботаник К. Раункиер рассматривал жизненные формы как результат приспособления растений к внешней среде. Решающее значение он придавал климату. В основу выделения жизненных форм Раункиер положил различия в приспособлении растений к переживанию неблагоприятного времени года. Эта приспособленность отражена в размещении почек или верхушек побегов по отношению к поверхности почвы (рис. 69). Большой вклад в разработку учения о жизненных формах внесли и другие зарубежные и российские ученые. Как синонимы и близкие понятия в экологии растений употребляются термины «растительные формы», «формы роста», «биологический тип», «экобиоморфа», «эпиморфа».
Рис. 69. Жизненные формы растений (по К. Раункиеру, 1907): 1–3– фанерофиты; 4–5 – хамефиты; 6–7 – гемикриптофиты; 8-11 – криптофиты; 12 – терофиты; 12а – семя с зародышем
Учение о жизненных формах особенно обогатили исследования И. Г. Серебрякова. Он дает следующее развернутое определение: «Жизненную форму у высших растений с эколого-морфологической точки зрения можно определить как своеобразный общий облик (габитус) определенной группы растений (включая их подземные органы), возникающий в их онтогенезе в результате роста и развития в определенных условиях среды. Исторически этот габитус развился в данных почвенно-климатических условиях как выражение приспособленности растений к этим условиям» (И. Г. Серебряков, 1964). По И. Г. Серебрякову, жизненную форму растения создает система его вегетативных органов. Жизненная форма – категория морфологическая и экологическая. Классификации жизненных форм растений. Таких классификаций к настоящему времени было создано множество. Современными исследователями чаще других используются системы К. Раункиера и И. Г. Серебрякова. Система К. Раункиера К. Раункиер использовал для классификации жизненных форм растений единственный, но имеющий большое приспособительное значение признак – положение почек возобновления по отношению к поверхности почвы. Сначала он разработал эту систему для растений Средней Европы, но затем распространил на растения всех климатических поясов. Все растения Раункиер разделил на пять типов (1903), в которых позднее выделил подтипы (1907). 1. Фанерофиты. Почки возобновления или верхушки побегов расположены в течение неблагоприятного времени года более или менее высоко в воздухе и подвергаются всем превратностям погоды. Подразделяются на 15 подтипов по высоте растений, по ритму развития листвы, по степени защищенности почек, по консистенции стебля. Один из подтипов – эпифитные фанерофиты. 2. Хамефиты. Почки возобновления у поверхности почвы или не выше 20–30 см. Зимой прикрыты снежным покровом. Подразделяются на 4 подтипа. 3. Гемикриптофиты. Почки возобновления или верхушки побегов на поверхности почвы, часто прикрыты подстилкой. Включают три подтипа и более мелкие подразделения. 4. Криптофиты. Почки возобновления или верхушки побегов сохраняются в почве (геофиты) или под водой (гелофиты и гидрофиты). Подразделяются на 7 подтипов. 5. Терофиты. Переносят неблагоприятное время года только в семенах. Раункиер считал, что жизненные формы складываются исторически как результат приспособления растений к климатическим условиям. Процентное распределение видов по жизненным формам в растительных сообществах на изучаемой территории он назвал биологическим спектром. Для разных зон и стран были составлены биологические спектры, которые могли служить индикаторами климата. Так, жаркий и влажный климат тропиков был назван «климатом фанерофитов», умеренно-холодные области имеют «климат гемикриптофитов», полярные страны – «климат хамефитов». Критики воззрений Раункиера отмечают, что его типы жизненных форм слишком обширны и неоднородны: хамефиты включают растения с разным отношением к климату, их много как в тундрах, так и в полупустынях. И не только современный климат определяет спектр жизненных форм, но и комплекс почвенно-литологических условий, а также история формирования флоры и влияние человеческой культуры. Тем не менее классификация жизненных форм растений по Раункиеру остается популярной и продолжает модифицироваться. Система И. Г. Серебрякова Наиболее разработанной классификацией жизненных форм покрытосеменных и хвойных на основе эколого-морфологических признаков является система И. Г. Серебрякова (1962, 1964). Она иерархична, в ней использована совокупность большого числа признаков в соподчиненной системе и приняты следующие единицы: отделы, типы, классы, подклассы, группы, подгруппы, иногда секции и собственно жизненные формы. Собственно жизненная форма является основной единицей экологической системы растений. Под жизненной формой как единицей экологической классификации И. Г. Серебряков понимает совокупность взрослых генеративных особей данного вида в определенных условиях произрастания, обладающих своеобразным обликом, включая надземные и подземные органы. Им выделены 4 отдела жизненных форм. 1. Отдел А. Древесные растения. Включает 3 типа: деревья, кустарники, кустарнички. 2. Отдел Б. Полудревесные растения. Включает 2 типа – полукустарники и полукустарнички. 3. Отдел В. Наземные травы. Включает 2 типа: поликарпические и монокарпические травы. 4. Отдел Г. Водные травы. Включает 2 типа: земноводные травы, плавающие и подводные травы. Выделение отделов основано на степени одревеснения надземных осей (древесные, полудревесные и травянистые растения), выделение типов – на относительной длительности жизни надземных осей или растений в целом. Классы в пределах типов выделяются на основании структуры побегов (лиановидные, ползучие, суккулентные и проч.), на основе специфики питания (сапрофиты и паразиты) или образа жизни (эпифиты). При характеристике собственно жизненной формы растений учитывается характер надземных побегов (удлиненные, укороченные, сильно ветвящиеся и образующие подушки, ползучие и т. п.), тип корневой системы (стержнекорневые, кистекорневые, корнеотпрысковые растения и т. п.), подземные побеги (короткие и длинные корневища, клубни, луковицы, столоны, каудексы и т. п.). Учитывается также общая длительность жизни и способность к повторному цветению (монокарпики и поликарпики) и др. Рассмотрим положение конкретных растений в системе жизненных форм И. Г. Серебрякова. Липа сердцевидная относится к отделу древесных растений, классу кронообразующих с полностью одревесневшими удлиненными побегами, подклассу наземных, группе с подземными корнями, подгруппе прямостоячих, секции одноствольных (лесного типа), к листопадным деревьям. Земляника лесная относится к отделу наземных трав, типу поликарпиков, классу травянистых поликарпиков с ассимилирующими побегами несуккулентного типа, подклассу столонообразующих и ползучих, группе столонообразующих, подгруппе наземностолонных. Собственную жизненную форму земляники лесной можно характеризовать как короткокорневищное кистекорневое растение с розеточными побегами и надземными столонами. И. Г. Серебряков отмечал недоработанность и незавершенность своей классификации вследствие слабой изученности жизненных форм растений разных сообществ, особенно дождевых тропических лесов. Габитус тропических деревьев определяется зачастую не только характером стволов и крон, но и корневыми системами, поэтому последние служат важным признаком при классификации жизненных форм деревьев. Травянистые растения имеют более короткую длительность надземных осей, разнообразные ритмы сезонного развития, разный характер надземных и подземных органов. Они часто вегетативно подвижны, обладают большой семенной продуктивностью, лучше деревьев приспособлены к освоению самых разнообразных местообитаний, иногда в очень суровых условиях. Поэтому разнообразие жизненных форм у наземных травянистых растений необыкновенно велико. Разнообразие и изменчивость жизненных форм растений. И. Г. Серебряков наметил параллельные ряды жизненных форм покрытосеменных растений и предполагаемые связи между ними (рис. 70). В сходных условиях как среди древесных, так и травянистых растений конвергентно возникли лиановидные, подушковидные, стелющиеся и суккулентные формы. Например, подушковидные формы древесные и травянистые часто встречаются в условиях хорошего освещения, но при низких температурах воздуха и почвы, при крайней сухости почвы и низкой влажности воздуха, при частых и сильных ветрах. Они обычны в высокогорьях, тундрах, пустынях, на субантарктических островах и в других местах с подобным комплексом условий.
Рис. 70. Параллельные ряды жизненных форм покрытосеменных растений и их предполагаемые связи (по И. Г. Серебрякову, 1955)
Сходные жизненные формы возникли конвергентно в разных систематических группах. Например, в условиях аридного климата пустынь одна и та же жизненная форма стеблевых суккулентов у кактусов в Америке, у молочаев и стапелий в Африке. Одну жизненную форму могут иметь как близкородственные виды (например, у манжеток), так и виды из разных семейств. К жизненной форме рыхлокустовых дерновинных поликарпиков с мочковатой корневой системой можно отнести овсяницу луговую и тимофеевку луговую (злаки), ожику волосистую (ситниковые), осоку заячью (осоковые) и т. д. В то же время один вид может иметь разные жизненные формы. Смена жизненных форм происходит у большинства растений в онтогенезе, так как по мере роста и развития габитус меняется иногда весьма существенно. У трав стержневая корневая система часто сменяется мочковатой, розеточные побеги – полурозеточными, каудекс из одноглавого превращается в многоглавый и т. д. Иногда габитус растения закономерно меняется по сезонам. У мать-и-мачехи и медуницы неясной весной от корневищ отходят удлиненные, с небольшими листьями генеративные побеги. В конце мая – начале июня после плодоношения они отмирают, а из почек на корневищах у этих же особей вырастают укороченные розеточные вегетативные побеги с крупными листьями, фотосинтезирующими до осени. У безвременника великолепного каждую осень генеративное растение представлено клубнелуковицей и отходящим от нее цветком, а весной – олиственным побегом, на верхушке которого созревает плод коробочка. В подобных случаях можно говорить о пульсирующих жизненных формах. Жизненная форма вида может меняться в пределах ареала в разных географических и экологических условиях. Многие виды деревьев на границах ареала образуют кустарниковые, нередко стелющиеся формы, например, ель обыкновенная на Крайнем Севере, ель сибирская на Южном Урале и в Хибинах. Отдельные виды деревьев представлены разными жизненными формами в одних и тех же географических районах и даже в одних и тех же фитоценозах (рис. 71). Например, липа может быть представлена в фитоценозах: 1) одноствольным деревом; 2) порослеобразующим деревом; 3) немногоствольным деревом с 2–3 стволами; 4) многоствольным деревом – так называемое дерево-куст; 5) куртинообразующим деревом; 6) торчками одноствольными; 7) торчками многоствольными; 8) факультативным стлаником. В центре ареала в оптимальных условиях – на Украине, в Тульской и Пензенской областях преобладают компактные жизненные формы липы, близ северо-восточной границы на Среднем Урале – стланиковые. Деревья-кусты появляются после рубки одноствольных деревьев и при повреждении главной оси морозами, вредителями. Факультативный стланик входит в подлесок, приурочен обычно к сильно затененным участкам, склонам и днищам оврагов. При улучшении световой обстановки стланик может перейти в кустовидную форму или стать куртинообразующим деревом. Куртина – это заросль, образовавшаяся из одного растения. Торчки – это угнетенные низкорослые растения, выросшие при недостатке света и влаги. У молодых растений отмирают верхушки лидирующих побегов, а затем и боковых. Прожив в таком состоянии 20–30 лет, торчки могут отмереть, так и не выйдя из травяного яруса, если условия освещения улучшаются, торчки могут образовать порослевые деревья. Другие деревья – ильм, клен, граб, черемуха и некоторые кустарники – бересклеты, жимолость, свидина, лещина и другие, также имеют широкий спектр жизненных форм. В лесах Дальнего Востока лимонник китайский в разных экологических условиях растет то как лиана, то как наземный кустарник. У травянистых растений тоже часто наблюдается внутривидовое разнообразие жизненных форм.
Рис. 71. Варианты жизненной формы у липы сердцевидной (по А. А. Чистяковой, 1978): 1– одноствольное дерево; 2 – порослеобразующее дерево; 3 – немногоствольное; 4 – многоствольное; 5 – куртинообразующее дерево; 6 – одноствольный торчок; 7 – многоствольный торчок; 8 – факультативный стланик
Жизненные формы животных Жизненные формы возникают как конвергентное сходство по многим признакам, составляющим в сумме общий габитус организмов, их внешний облик. Классификации жизненных форм животных, как и растений, весьма разнообразны и зависят от принципов, которые кладут в их основу. На морфологии млекопитающих, по-видимому, в наибольшей степени сказывается характер их передвижения в разных средах. А. Н. Формозов выделил на этом основании среди зверей следующие адаптивные типы: 1) наземные формы; 2) подземные (землерои); 3) древесные; 4) воздушные и 5) водные. Между типами есть переходы. В пределах каждой группы особенности поступательного движения и образа жизни формируют более специфические приспособительные формы. Так, наземные млекопитающие передвигаются в основном посредством ходьбы, бега и прыжков, что проявляется в их внешнем облике. Например, прыгающие животные: кенгуру, тушканчики, прыгунчики, кенгуровые крысы – отличаются компактным телом с удлиненными задними конечностями и значительно укороченными передними. Очень сильно развиты мышцы – разгибатели спины, увеличивающие мощность толчка. Длинный хвост играет роль балансира и руля, позволяющего резко изменять направление движения. Он служит также дополнительной точкой опоры. Сходный тип передвижения и внешний облик имели и некоторые вымершие динозавры мезозоя. Естественно, что, кроме общих адаптивных признаков, каждый вид отличается деталями внешнего строения в зависимости от экологической специфики. Жизненные формы отчетливо выделяются в пределах любой крупной таксономической группы животных, характеризующейся экологическим разнообразием видов (рис. 72, 73). Во внешнем облике птиц в наибольшей мере проявляется приуроченность их к определенным типам местообитаний и характер передвижения при добывании пищи. В связи с этим различают птиц: 1) древесной растительности; 2) открытых пространств суши; 3) болот и отмелей; 4) водных пространств. В каждой из указанных групп выделяют специфические формы: а) добывающие пищу посредством лазания (многие голуби, попугаи, кукушки, дятловые, воробьиные); б) добывающие пищу в полете (в лесах – совы, козодои и др., на открытых пространствах – ржанковые, длиннокрылые, над водой – трубконосые и т. п.); в) кормящиеся при передвижении по земле (лесные – большинство куриных, казуары, киви и др., на открытых пространствах – страусы, нанду, журавлиные, на болотах и отмелях – голенастые, фламинго, некоторые воробьиные); г) добывающие пищу с помощью плавания и ныряния (пингвины, гагары, поганки, ряд трубконосых, большинство веслоногих и гусиных и др.).
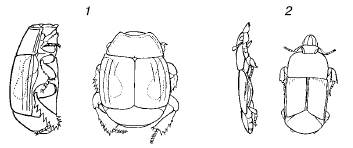
Рис. 72. Форма тела жуков-карапузиков из различных местообитаний (вид сбоку и сверху, по W. Kuhnelt, 1970): 1– из навоза; 2 – из-под коры деревьев
Рис. 73. Формы тела брюхоногих моллюсков, ведущих разный образ жизни
Жизненные формы таких мелких почвенных членистоногих, как коллемболы, выделяют на основе приуроченности их к определенным слоям почвенного профиля, так как в почве с глубиной резко меняется весь комплекс условий обитания: размеры полостей, освещенность, режим температуры и влажности и т. п. Атмобионты – виды, населяющие верхние слои подстилки и способные подниматься на нижние части растений, характеризуются более крупными размерами, полностью развитыми глазами, относительно длинными придатками (антеннами, ногами и прыгательной вилкой), хорошо выраженной пигментацией. Эуэдафические виды – обитатели минеральных тонкопорозных слоев почвы – отличаются мелкими размерами, сильным укорочением придатков, редукцией вилки, полным отсутствием глаз и пигмента. Гемиэдафические формы характеризуются промежуточными чертами: частичной редукцией глаз, рассеянным пигментом, укороченной вилкой и др. (рис. 74).
Рис. 74. Жизненные формы коллембол разных семейств (М. Б. Потапов)
Среди саранчовых по форме (конфигурация тела, структура головы, строение конечностей, летательного аппарата, тип покровительственной окраски) хорошо различаются тамнобионты– обитатели кустарников и деревьев; населяющие травянистый ярус хортобионты; жители надпочвенного слоя органических остатков герпетобионты; обитатели открытых участков грунта: эремобионты – на поверхности плотных глинистых почв, псаммобионты – на песках и петробионты – на каменистых участках с разреженной растительностью. Сходные жизненные формы встречаются в сходных условиях жизни в разных зоогеографических областях и на разных материках (например, тушканчики Евразии, кенгуровые крысы Австралии, прыгунчики Африки). Жизненная форма наглядно свидетельствует об образе жизни вида. Форма тела рыб, например, связана с тем, каким способом и с какой скоростью они передвигаются в воде – более плотной и вязкой среде, чем воздух. Торпедовидный тип тела имеют самые хорошие пловцы пелагиали (скумбрия, кефаль, лососи и др.). Виды со стреловидной, более вытянутой формой также приближаются к ним по скорости плавания в толще вод (сарган, щука). Сплющенное с боков тело – у рыб с меньшей скоростью, но такая форма позволяет хорошо маневрировать. Рыбы со змеевидным телом (угри, морские иглы и др.) обитают в зарослях. Морские коньки и тряпичники имитируют водоросли по форме. Лентовидное тело – сильно вытянутое и сплющенное с боков – у плохих пловцов (сельдяной король и т. п.). Рыбы, плавающие в придонных слоях, могут быть также шаровидными (кузовки), сплющенными дорзовентрально (скаты, морской черт). Движение рыб обеспечивается либо изгибанием всего тела, либо, при неподвижном теле, колебательными движениями разных плавников. Глава 7. Биоценозы Понятие о биоценозе Каждый организм живет в окружении множества других, вступает с ними в самые разнообразные отношения как с отрицательными, так и с положительными для себя последствиями и в конечном счете не может существовать без этого живого окружения. Связь с другими организмами – необходимое условие питания и размножения, возможность защиты, смягчения неблагоприятных условий среды, а с другой стороны – это опасность ущерба и часто даже непосредственная угроза существованию индивидуума. Всю сумму воздействий, которые оказывают друг на друга живые существа, объединяют названием биотические факторы среды. Непосредственное живое окружение организма составляет его биоценотическую среду. Представители каждого вида способны существовать лишь в таком живом окружении, где связи с другими видами обеспечивают им нормальные условия жизни. Иными словами, многообразные живые организмы встречаются на Земле не в любом сочетании, а образуют определенные сожительства, или сообщества, в которые входят виды, приспособленные к совместному обитанию. Группировки совместно обитающих и взаимно связанных видов называют биоценозами (от лат. «биос» – жизнь, «ценоз» – общий). Приспособленность членов биоценоза к совместной жизни выражается в определенном сходстве требований к важнейшим абиотическим условиям среды и закономерных отношениях друг с другом. Понятие «биоценоз» – одно из важнейших в экологии. Этот термин был предложен в 1877 г. немецким гидробиологом К. Мёбиусом, изучавшим места обитания устриц в Северном море. Он установил, что устрицы могут жить лишь в определенных условиях (глубина, течения, характер грунта, температура воды, соленость и т. п.) и что вместе с ними постоянно обитает определенный набор других видов – моллюсков, рыб, ракообразных, иглокожих, червей, кишечнополостных, губок и т. п. (рис. 75). Все они связаны между собой и подвержены влиянию окружающих условий. Мёбиус обратил внимание на закономерность такого сожительства. «Наука, однако, не имеет слова, которым такое сообщество живых существ могло бы быть обозначено, – писал он. – Нет слова для обозначения сообщества, в котором сумма видов и особей, постоянно ограничиваемая и подвергающаяся отбору под влиянием внешних условий жизни благодаря размножению непрерывно владеет некоторой определенной территорией. Я предлагаю термин «биоценоз» для такого сообщества. Всякое изменение в каком-либо из факторов биоценоза вызывает изменения в других факторах последнего». По Мёбиусу, возможность видов длительно сосуществовать друг с другом в одном биоценозе представляет результат естественного отбора и сложилась в историческом развитии видов. Дальнейшее изучение закономерностей сложения и развития биоценозов привело к возникновению особого раздела общей экологии – биоценологии. Масштабы биоценотических группировок организмов очень различны, от сообществ подушек лишайников на стволах деревьев или разлагающегося пня до населения целых ландшафтов: лесов, степей, пустынь и т. п.
Рис. 75. Биоценозы Черного моря (по С. А. Зернову, 1949): А – биоценоз скал: 1– краб Pachygrapsis; 2 – усоногие рачки Balanus; 3 – моллюск Patella; 4–5 – водоросли; 6 – мидии; 7 – актинии; 8 – морской ерш; Б – биоценоз песка: 9– немеретины; 10 – черви Saccocirrus; 11 – бокоплавы; 12 – моллюски Venus; 13 – рыбы-султанки; 14 – камбалы; 15 – раки-отшельники; В – биоценоз зарослей зостеры: 16– зостера; 17 – морские иглы; 18 – зеленушки; 19 – морские коньки; 20 – креветки; Г – биоценоз устричника: 21– устрицы; 22 – морские гребешки; Д – биоценоз мидиевого ила: 23– мидии; 24 – красная водоросль; 25 – красная губка Suberites; 26 – асцидия Ciona; Е – биоценоз фазеолинового ила: 27– моллюск фазеолина; 28 – иглокожие амфиуры; 29 – моллюск Trophonopsis; Ж – сероводородное царство бактерий; З – биоценоз планктона открытого моря: 31– медуза и др.
Термин «биоценоз» в современной экологической литературе чаще употребляют применительно к населению территориальных участков, которые на суше выделяют по относительно однородной растительности (обычно по границам растительных ассоциаций), например биоценоз ельника-кисличника, биоценоз суходольного луга, сосняка-беломошника, биоценоз ковыльной степи, пшеничного поля и т. д. При этом имеется в виду вся совокупность живых существ – растений, животных, микроорганизмов, приспособленных к совместному обитанию на данной территории. В водной среде различают биоценозы, соответствующие экологическим подразделениям частей водоемов, например биоценозы прибрежных галечных, песчаных или илистых грунтов, абиссальных глубин, пелагические биоценозы крупных круговоротов водных масс и т. п. По отношению к более мелким сообществам (населению стволов или листвы деревьев, моховых кочек на болотах, нор, муравейников, разлагающихся пней и т. д.) применяют разнообразные термины: «микросообщества», «биоценотические группировки», «биоценотические комплексы» и др. Принципиальной разницы между биоценотическими группировками разных масштабов нет. Более мелкие сообщества входят составной, хотя и относительно автономной частью в более крупные, а те, в свою очередь, являются частями сообществ еще больших масштабов. Так, все живое население моховых и лишайниковых подушек на стволе дерева – это часть более крупного сообщества организмов, связанного с данным деревом и включающего его подкоровых и наствольных обитателей, население кроны, ризосферы и т. п. В свою очередь, эта группировка – лишь одна из составных частей лесного биоценоза. Последний входит в более сложные комплексы, образующие в конечном счете весь живой покров Земли. Таким образом, организация жизни на биоценотическом уровне иерархична. С увеличением масштабов сообществ усиливается их сложность и доля непрямых, косвенных связей между видами. Естественные объединения живых существ имеют собственные законы сложения, функционирования и развития, т. е. представляют собой природные системы. Обсуждая общие принципы организации жизни на Земле, известный отечественный биолог В. Н. Беклемишев писал: «Все биоценотические ступени организации, от океанических и эпиконтинентальных комплексов до каких-нибудь микроскопических лишайников на стволе сосны – очень мало индивидуализированы, мало интегрированы, низко организованы, слабо замкнуты. Это расплывчатые, не очень определенные, часто трудно уловимые коллективные образования, сложно переплетенные между собой, незаметно переходящие друг в друга и тем не менее вполне реальные, существующие и действующие, которые нам надо понимать во всей их сложности и расплывчатости, что и составляет задачу биоценологии со всеми ее ответвлениями». Таким образом, являясь, как и организмы, структурными единицами живой природы, биоценозы тем не менее складываются и поддерживают свою устойчивость на основе иных принципов. Они представляют собой системы так называемого каркасного типа, без особых управляющих и координирующих центров (как, например, нервная или гуморальная системы организмов), но также строятся на многочисленных и сложных внутренних связях, имеют закономерную структуру и определенные границы устойчивости. Важнейшими особенностями систем, относящихся к надорганизменному уровню организации жизни, по классификации немецкого эколога В. Тишлера, являются следующие: 1. Сообщества всегда возникают, складываются из готовых частей (представителей различных видов или целых комплексов видов), имеющихся в окружающей среде. Этим способ их возникновения отличается от формирования отдельного организма, особи, которое происходит путем постепенного дифференцирования зачатков. 2. Части сообщества заменяемы. Один вид (или комплекс видов) может занять место другого со сходными экологическими требованиями без ущерба для всей системы. Части же (органы) любого организма уникальны. 3. Если в целостном организме поддерживается постоянная координация, согласованность деятельности его органов, клеток и тканей, то надорганизменная система существует в основном за счет уравновешивания противоположно направленных сил. Интересы многих видов в биоценозе прямо противоположны. Например, хищники – антагонисты своих жертв, но тем не менее они существуют вместе, в рамках единого сообщества. 4. Сообщества основаны на количественной регуляции численности одних видов другими. 5. Предельные размеры организма ограничены его внутренней наследственной программой. Размеры надорганизменных систем определяются внешними причинами. Так, биоценоз сосняка-беломошника может занимать небольшой участок среди болот, а может простираться на значительное расстояние на территории с относительно однородными абиотическими условиями. |
Последнее изменение этой страницы: 2017-04-13; Просмотров: 415; Нарушение авторского права страницы