|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Структура и регуляция генов прокариотСтр 1 из 3Следующая ⇒
Тонкая структура генов Существование дискретных наследственных факторов в половых клетках было гипотетически постулировано Г. Менделем в 1865 г. С начала формирования хромосомной теории считалось, что эти факторы являются не только единицами наследственности, но и конкретными физическими величинами, связанными с хромосомами. Для их обозначение датский генетик В. Иогансен в 1909 г. ввел термин «гены». Первоначально, под геном понималась последовательность нуклеотидов, кодирующая синтез отдельной полипептидной цепи. Это представление было основано на концепции Дж. Бидла и Э. Тэйтема, сформулированное в 40-е г., согласно которому всякий ген определяет синтез одного фермента. Эволюция этого положения выглядит так: «один ген- один фермент» → «один ген – один белок» → «один ген – один полипепдид». Дальнейшие исследования структуры гена, связанные с расшифровкой генетического кода и экспериментами по изучению мутаций у различных организмов привели к пониманию того, что ген не является неделимой единицей, и мутации могут происходить и внутри гена. Больший вклад в понимание структуры гена внесли работы Маршалла У. Ниренберга по расшифровке генетического кода и его функционирования в синтезе белков, Северо Очоа, изучавшего механизмы биологического синтеза рибонуклеиновой и дезоксирибонуклеиновой кислот, а также эксперименты Сеймура Бензера и Френсиса Крика, которые в 1961 г. установили, что генетический код триплетен. Схема проведенного эксперимента следующая - проводили добавление или удаление одного нуклеотида в известную последовательность: " +" мутация - вставка одного нуклеотида. " -" мутация - выпадение одного нуклеотида. В ходе эксперимента выяснили, что: Одиночная " +" или " -" мутация в начале гена портит весь ген. Двойная " +" или " -" мутация тоже портит весь ген. Тройная " +" или " -" мутация в начале гена портит лишь его часть. Четверная " +" или " -" мутация опять портит весь ген. Этот эксперимент доказал, что код триплетен и внутри гена нет знаков препинания. Эксперимент был проведен на двух рядом расположенных фаговых генах и показал, кроме того, сто между соседними генами присутствуют знаки препинания. Дальнейшее изучение структуры генов привело к формированию понятия «генетический код» и определению общих свойств генетического кода. Под кодом ДНК, или генетическим кодом понимается отражение структуры белков в виде триплетов ДНК. Благодаря генетическому коду устанавливается однозначное соответствие между нуклеотидными последовательностями нуклеиновых кислот и аминокислотами, входящими в состав белков. Генетический код обладает следующими основными свойствами: 1.Генетический код триплетен: каждая аминокислота кодируется триплетом нуклеотидов ДНК и соответствующим триплетом иРНК. При этом кодоны ничем не отделены друг от друга (отсутствуют «запятые»). 2.Генетический код является избыточным (вырожденным): почти все аминокислоты могут кодироваться разными кодонами. Только двум аминокислотам соответствует по одному кодону: метионину (АУГ) и триптофану (УГГ). Зато лейцину, серину и аргинину соответствует по 6 разных кодонов. 3.Генетический код является неперекрывающимся: каждая пара нуклеотидов принадлежит только одному кодону (исключения обнаружены у вирусов). 4. Генетический код един для подавляющего большинства биологических систем. Однако имеются и исключения, например, у инфузорий и в митохондриях разных организмов. Поэтому генетический код называют квазиуниверсальным. 5. Компактность, или отсутствие внутригенных знаков препинания. Внутри гена каждый нуклеотид входит в состав значащего кодона.
В классической генетике ген определяется не только как физическая последовательность нуклеотидов, но и как единица генетического материала, обладающая определенными свойствами: функции, мутации, рекомбинации. За время исследований, полученные данные привели к изменению первоначальных представлений о строении и свойствах генов. Изначально предполагалось, что ген - это функциональная единица, то есть нечто, определяющее отдельный признак. Такое представление сохранилось и до сих пор, но сейчас уже известно, что на один и тот же признак могут воздействовать различные гены и что при мутации гены могут давать один и тот же фенотип. Кроме того, ген определяли как единицу мутации. Эксперименты Бензера показали, что так, как ген представляет собой линейную последовательность многих участков, то мутации могут затрагивать каждый их этих составляющих. Гены изначально определялись и как единицы рекомбинаций, хотя сейчас известно, что гены не представляют собой неделимые «бусины» на цепи, а рекомбинации происходят и внутри генов. Это и следовало ожидать, если предположить, что ген представляет собой всего лишь участок ДНК, любые нуклеотидные пары которой могут изменяться, в результате мутации и рекомбинаций. Структура генов эукариот Позже, когда исследователи начали изучать гены различных белков в клетках эукариот, обнаружилось, что взаимодействие генов и белков в этих организмах более сложное, чем взаимодействие генов и белков прокариот. Первые примеры такого взаимодействия были получены в 1977 году в лабораториях Филиппа Шарпа и Пьера Шамбона. Вместе со своими коллегами они гибридизировали мРНК различных генов с теми ДНК, с которых были сняты эти информационные копии. У бактерий последовательность мРНК идентична последовательности кодирующей цепи ДНК (за исключением того, что место тимина занимает урацил), поэтому структура гибридных молекул была достаточно проста. Но когда под электронным микроскопом были сделаны снимки гибридных молекул генов эукариот, то в них обнаружился ряд петель. Это значит, что мРНК и ДНК имеют не совсем идентичную последовательность, и петли были как раз теми местами, в которых они не могли соединяться. Когда последовательность мРНК сравнили с последовательностью ДНК, стало понятно, что кодирующая последовательность генов в некоторых местах прерывается некодирующей последовательностью, то есть некоторые нуклеотиды не кодируют синтез белка. Впоследствии выяснилось, что это типичная картина для ДНК эукариот. Кодирующая последовательность гена называется экзоном, а некодирующая последовательность — интроном. Некоторые гены имеют в своей структуре несколько интронов. Часто обнаруживают и такие гены, в которых больше интронов, чем экзонов.
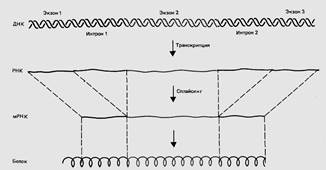
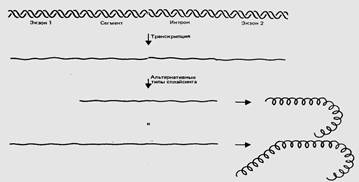
Рис. 2. Прерывистые гены экспрессируются в виде РНК- предшественника, из которой затем удаляются интроны; экзоны, соединяясь вместе, образуют мРНК. В общем случае при транскрипции генов эукариот образуются большие молекулы РНК, содержащие как экзоны, так и интроны. Таким образом, полученные транскрипты, являются гетерогенными, т.е. они не на всем протяжении способны к трансляции. Поэтому первичные транскриптоны эукариот называются гетерогенной ядерной РНК (гяРНК) или про-мРНК. В дальнейшем происходит преобразование про-мРНК, или ее «созревание», при котором она еще в ядре вовлекается в процессинг, состоящий в том, чтоособые комплексы ферментов (сплайсингсомы) вырезают из транскрипта все интроны. Затем экзоны соединяют в одну мРНК, кодирующую производство белка. Процесс соединения экзонов называется сплайсингом (сращивание). В результате процессинга образуется «зрелая» мРНК, которая по размерам значительно короче, чем про-РНК. Молекулы про-мРНК содержат, в среднем, более 50 000 нуклеотидов, а созревшая мРНК – 500-3000 нуклеотидов. Далее полученная м-РНК транслируется как обычно. Сплайсинг характерен для эукариот и архебактерий, но иногда встречается и у прокариот. Причины, по которым природа придерживается такой структуры, до сих пор не ясны, но ее можно объяснить как с эволюционной точки зрения, так и с точки зрения развития организма. Если говорить об эволюции, то такая структура ценна тем, что позволяет экспериментировать с генами и создавать новые гены. Кроссинговер может происходить внутри интронов, и в таком случае ошибки будут несущественными, а при рекомбинации могут образоваться новые экзоны и как следствие новые белки. Часто бывает так, что отдельный экзон кодирует отдельную область, или домен, белка, то есть отдельную часть белка с особыми функциями. Поэтому включение в ген нового экзона приведет к созданию белка с новыми областями и, возможно, с новыми функциями. Такое изменение генетической структуры может служить источником эволюции. Другой особенностью работы генов эукариот является то, что сплайсинг мРНК, полученной с одного и того же локуса ДНК, может происходить по-разному. Это явление получило название альтернативный сплайсинг и оно приводит к тому, что конечный продукт –«созревшая» мРНК имеет разную нуклеотидную последовательность. Сущность альтернативного сплайсинга заключается в том, что одни и те же участки исходной мРНК могут быть и интронами, и экзонами. Тогда одному и тому же участку ДНК соответствует несколько типов зрелой мРНК и, соответственно, несколько разных форм одного и того же белка. На рис.3 представлен вариант альтернативного сплайсинга участков гена эукариот. В первом случае экзон 1 и экзон 2 соединяются вместе, а участки, обозначенные «сегмент» и «интрон», удаляются из РНК. Во втором случае соединяются экзон 1, сегмент и экзон 2, а удаляется только интрон. Таким образом, область, обозначенная «сегмент», служит интроном в первом случае и экзоном - во втором. В результате этих двух типов сплайсинга образуются два белка, которые имеют одинаковые концы, но различаются по средней части. Так одна последовательность ДНК может кодировать более одного белка.
Рис 3. Гены, имеющие альтернативные пути экспрессии, бывает трудно разграничить.
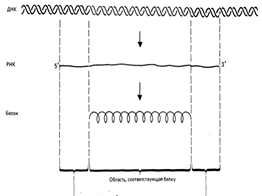
Существует также транс–сплайсинг, который заключается в соединение экзонов, кодируемых разными генами (иногда даже из разных хромосом), в одну зрелую молекулу мРНК. Таким образом, структура интрон-экзон позволяет одной нуклеотидной последовательности кодировать синтез более одного белка. Гены эукариот, в силу их более сложной организации и функционирования, в результате чего получаются разные продукты-полипептиды, получили название «транскриптоны». Структура гена в полной мере проявляется в свойствах белков. Это конечное проявление структуры генов -синтез полипептидов по созданной мРНК, называется проявлением генов. Этапы биосинтеза белков Генетическая информация о структуре белка хранится в виде последовательности триплетов ДНК. При этом лишь одна из цепей ДНК служит матрицей для транскрипции. Биосинтез белков в клетках представляет собой последовательность реакций матричного типа, в ходе которых последовательная передача наследственной информации с одного типа молекул на другой приводит к образованию полипептидов с генетически обусловленной структурой. Биосинтез белков представляет собой начальный этап реализации, или экспрессии генетической информации. К главным матричным процессам, обеспечивающим биосинтез белков, относятся транскрипция ДНК и трансляция мРНК. Транскрипция ДНК заключается в переписывании информации с ДНК на мРНК (матричную, или информационную РНК). Трансляция мРНК заключается в переносе информации с мРНК на полипептид. Копирование мРНКначинается с прикрепления РНК-полимеразы к участку ДНК, который называется промотором. Однако, учитывая сведения о возможности альтернативного сплайсинга, возможны случаи, когда гены, даже, расположенные рядом, будут транскрибироваться с разных цепей. Таким образом, для транскрипции могут использоваться обе цепи ДНК. При транскрипции комплементарных цепей ДНК используются разные РНК-полимеразы, а направление их движения по цепи определяется последовательностью промотора. Так как цепи ДНК инвертированы относительно друг друга, а синтез мРНК, также, как синтез ДНК идет только в направлении от 5ꞌ к 3ꞌ концу, то и транскрипции на ДНК идут в противоположных направлениях. Цепь ДНК, которая содержит те же последовательности, что и мРНК, называется кодирующей, а цепь, обеспечивающая синтез мРНК (на основе комплементарного спаривания) – антикодирующей. Антикодирующая цепь также называется транскрибируемой. Кроме мРНК в клетке образуются и другие продукты транскрипции ДНК. К ним относятся молекулы рРНК и тРНК, которые также являются участниками синтеза полипептидов. Все эти РНК называются ядерными. Если рассматривать процентное содержание этих трех видов РНК в клетке, то на долю зрелой мРНК приходится около 5 % от общего содержания РНК, на долю тРНК – около 10 %, а большая часть – до 85 % приходится на рРНК. Все РНК транскрибируются с ДНК из рибонуклеотидтрифосфатов с освобождением пирофосфата при участии РНК-полимераз. У прокариот присутствует только один вид РНК-полимеразы, которая обеспечивает синтез мРНК, рРНК и тРНК. В клетках эукариот присутствует три вида РНК- полимераз (I, II, III). Каждая из этих РНК-полимераз, прикрепляясь к промотору на ДНК, обеспечивает транскрипцию разных последовательностей ДНК. РНК-полимераза I синтезирует крупные рРНК (основные молекулы РНК больших и малых субъединиц рибосом). РНК-полимераза II синтезирует все мРНК и часть малых рРНК, РНК-полимераза III синтезирует тРНК и РНК 5s –субъединиц рибосом. Для связывания РНК-полимераз с промотором необходимы особые белки, выполняющие функцию факторов инициации транскрипции (TF I, TF II, TF III для соответствующих полимераз). С учетом этих позиций, основные этапы биосинтеза белков состоят в следующем: 1 этап. Транскрипция ДНК. На транскрибируемой цепи ДНК с помощью ДНК-зависимой РНК-полимеразы достраивается комплементарная цепь мРНК. Молекула мРНК является точной копией нетранскрибируемой цепи ДНК с той разницей, что вместо дезоксирибонуклеотидов в ее состав входят рибонуклеотиды, в состав которых вместо тимина входит урацил. 2 этап. Процессинг (созревание) мРНК. Синтезированная молекула мРНК (первичный транскрипт) подвергается дополнительным превращениям. В большинстве случаев исходная молекула мРНК разрезается на отдельные фрагменты. Одни фрагменты – интроны – расщепляются до нуклеотидов, а другие – экзоны – сшиваются в зрелую мРНК. Все стадии процессинга мРНК происходят в РНП-частицах (рибонуклеопротеидных комплексах). По мере синтеза про-мРНК, она тут же образует комплексы с ядерными белками – информоферами и образует ядерные и цитоплазматические комплексы (мРНК плюс информоферы) - информосомы. Таким образом, мРНК не бывает свободной от белков. На всем пути следования до завершения трансляции мРНК защищена от нуклеаз. Кроме того, белки придают ей необходимую конформацию. 3 этап. Трансляция мРНК. Полученная при транскрипции молекула мРНК служит матрицей для синтеза полипептида на рибосомах. Триплеты мРНК, кодирующие определенную аминокислоту, называются кодоны. В трансляции принимают участие молекулы тРНК. Каждая молекула тРНК содержит антикодон – распознающий триплет, в котором последовательность нуклеотидов комплементарна по отношению к определенному кодону мРНК. Каждая молекула тРНК способна переносить строго определенную аминокислоту. Молекула тРНК по общей конформации напоминает клеверный лист на черешке. «Вершина листа» несет антикодон. Существует 61 тип тРНК с разными антикодонами. К «черешку листа» присоединяется аминокислота (существует 20 аминокислот, участвующих в синтезе полипептида на рибосомах). Каждой молекуле тРНК с определенным антикодоном соответствует строго определенная аминокислота. В то же время, определенной аминокислоте обычно соответствует несколько типов тРНК с разными антикодонами. Аминокислота ковалентно присоединяется к тРНК с помощью ферментов – аминоацил-тРНК-синтетаз. Эта реакция называется аминоацилированием тРНК. Соединение тРНК с аминокислотой называется аминоацил–тРНК.
Трансляция (как и все матричные процессы) включает три стадии: инициацию (начало), элонгацию (продолжение) и терминацию (окончание). Инициация. Сущность инициации заключается в образовании пептидной связи между двумя первыми аминокислотами полипептида. - Первоначально образуется инициирующий комплекс, в состав которого входят: малая субъединица рибосомы, специфические белки (факторы инициации) и специальная инициаторная метиониновая тРНК с аминокислотой метионином – Мет–тРНКМет. Инициирующий комплекс узнает начало мРНК, присоединяется к ней и скользит до точки инициации (начала) биосинтеза белка: в большинстве случаев это стартовый кодон АУГ. Между стартовым кодоном мРНК и антикодоном метиониновой тРНК происходит кодонзависимое связывание с образованием водородных связей. Затем происходит присоединение большой субъединицы рибосомы. - При объединении субъединиц образуется целостная рибосома, которая несет два активных центра (сайта): А–участок (аминоацильный, который служит для присоединения аминоацил-тРНК) и Р–участок (пептидилтрансферазный, который служит для образования пептидной связи между аминокислотами). Первоначально Мет–тРНКМет находится на А–участке, но затем перемещается на Р–участок. На освободившийся А–участок поступает аминоацил-тРНК с антикодоном, который комплементарен кодону мРНК, следующему за кодоном АУГ. Например, это Гли–тРНКГли с антикодоном ЦЦГ, который комплементарен кодону ГГЦ. В результате кодонзависимого связывания между кодоном мРНК и антикодоном аминоацил-тРНК образуются водородные связи. Таким образом, на рибосоме рядом оказываются две аминокислоты, между которыми образуется пептидная связь. Ковалентная связь между первой аминокислотой (метионином) и её тРНК разрывается. - После образования пептидной связи между двумя первыми аминокислотами рибосома сдвигается на один триплет. В результате происходит транслокация (перемещение) инициаторной метиониновой тРНКМет за пределы рибосомы. Водородная связь между стартовым кодоном и антикодоном инициаторной тРНК разрывается. В результате свободная тРНКМет отщепляется и уходит на поиск своей аминокислоты. При этом, вторая тРНК вместе с аминокислотой (Гли–тРНКГли) в результате транслокации оказывается на Р–участке, а А–участок освобождается. Элонгация. Сущность элонгации заключается в присоединении последующих аминокислот, то есть в наращивании полипептидной цепи. Рабочий цикл рибосомы в процессе элонгации состоит из трех шагов: кодонзависимого связывания мРНК и аминоацил-тРНК на А–участке, образования пептидной связи между аминокислотой и растущей полипептидной цепью и транслокации с освобождением А–участка. На освободившийся А–участок поступает аминоацил-тРНК с антикодоном, соответствующим следующему кодону мРНК (например, это Тир–тРНКТир с антикодоном АУА, который комплементарен кодону УАУ). На рибосоме рядом оказываются две аминокислоты, между которыми образуется пептидная связь. Связь между предыдущей аминокислотой и её тРНК (в нашем примере между глицином и тРНКГли) разрывается. Затем рибосома смещается еще на один триплет, и в результате транслокации тРНК, которая была на Р–участке (в нашем примере тРНКГли), оказывается за пределами рибосомы и отщепляется от мРНК. А–участок освобождается, и рабочий цикл рибосомы начинается сначала. Терминация. Заключается в окончании синтеза полипептидной цепи. Энергетика биосинтеза белков. Биосинтез белков – очень энергоемкий процесс. При аминоацилировании тРНК затрачивается энергия одной связи молекулы АТФ, при кодонзависимом связывании аминоацил-тРНК – энергия одной связи молекулы ГТФ, при перемещении рибосомы на один триплет – энергия одной связи еще одной молекулы ГТФ. В итоге на присоединение аминокислоты к полипептидной цепи затрачивается около 90 кДж/моль. При гидролизе же пептидной связи высвобождается лишь 2 кДж/моль. Таким образом, при биосинтезе большая часть энергии безвозвратно теряется (рассеивается в виде тепла).
Регуляция на генном уровне 1.1.Модификация ДНК. Замена мажорных «обычных» азотистых оснований – аденина, гуанина, цитозина и тимина – на минорные «редкие» азотистые основания, обычно на метил-цитозин или метил-гуанин. Доказано, что метилирование цитозина существенно влияет на экспрессию генов. Например, активные гены гемоглобина менее метилированы, чем неактивные. 1.2. Различные случаи программированных количественных изменений ДНК. Примером регуляции, обусловленной транспозицией, служит феномен смены фаз (типа жгутиков) у сальмонелл. Действующий в клетках сальмонелл переключатель содержит промотор, который может изменять свою пространственную ориентацию. В одной ориентации промотор обеспечивает транскрипцию гена Н2, кодирующего синтез жгутиков одного типа, с одновременной репрессией гена H1, кодирующего синтез жгутиков другого типа, и наоборот. 1.3. Сплайсинг ДНК. Регуляция, связанная со сплайсингом ДНК, изучена на примере генов, кодирующих синтез антител. Известно, что разнообразные чужеродные вещества – антигены, попадающие в наш организм, – связываются особыми белками – антителами, или иммуноглобулинами. Млекопитающие могут продуцировать до миллиона различных антител, которые вырабатываются Т- и В-лимфоцитами иммунной системы. Существует особый раздел генетики – иммуногенетика, – который изучает генетический контроль иммунного ответа. Основу молекул иммуноглобулинов составляет сложный белок, состоящий из четырех полипептидных цепей – двух тяжелых (Н) и двух легких (L), – связанных дисульфидными мостиками. Оба типа цепей имеют константные (С) и вариабельные (V) участки. Доказано, что у эмбрионов фрагменты ДНК, кодирующие V- и С-участки, пространственно разделены. При развитии системы иммунитета у позвоночных животных и человека происходит дифференцировка лимфоцитов, в ходе которой гены, кодирующие V- и С-участки, перестраиваются таким образом, что в итоге они оказываются частями одного и того же гена, транскрибируемого как целое. Таким образом, сплайсинг ДНК обеспечивает сшивание консервативных (т.е. постоянно присутствующих) районов этих генов с различными варьирующими. В результате появляется большое число типов антител, поскольку любая консервативная область может быть присоединена к любой варьирующей. 1.4. Диминуция хроматина. У некоторых организмов (у аскарид, циклопов) в соматических клетках происходит необратимая утрата части генетического материала (от 20 до 80% ДНК). В полном объеме исходная генетическая информация сохраняется только в клетках зародышевого пути, т. е. в клетках, которые дадут в дальнейшем начало половым клеткам. Именно гаметы содержат всю полноту генетической информации данного вида и составляют непрерывный, потенциально бессмертный зародышевый путь. А. Вайсман считал диминуцию хроматина универсальным механизмом дифференцировки клеток и тканей, однако в дальнейшем было показано, что этот способ дифференцировки встречается довольно редко. Например, подобное явление наблюдается у инфузорий: в диплоидном микронуклеусе полностью сохраняется исходный набор генов, а в полиплоидном макронуклеусе ~10% генов (правда, за счет полиплоидизации оставшаяся информация многократно дублируется). 1.5. Изменение активности целых хромосом. Известно, что у самок млекопитающих в кариотипе присутствует две X-хромосомы, а у самцов одна X- и одна Y-хромосома. Несмотря на то, что женские особи млекопитающих имеют две Х-хромосомы, а мужские – только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у самок в каждой клетке полностью инактивирована одна Х-хромосома. Эту хромосому можно видеть в интерфазе в форме гетерохроматинового тельца, названного тельцем Барра. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. При этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики. Тонкая структура генов Существование дискретных наследственных факторов в половых клетках было гипотетически постулировано Г. Менделем в 1865 г. С начала формирования хромосомной теории считалось, что эти факторы являются не только единицами наследственности, но и конкретными физическими величинами, связанными с хромосомами. Для их обозначение датский генетик В. Иогансен в 1909 г. ввел термин «гены». Первоначально, под геном понималась последовательность нуклеотидов, кодирующая синтез отдельной полипептидной цепи. Это представление было основано на концепции Дж. Бидла и Э. Тэйтема, сформулированное в 40-е г., согласно которому всякий ген определяет синтез одного фермента. Эволюция этого положения выглядит так: «один ген- один фермент» → «один ген – один белок» → «один ген – один полипепдид». Дальнейшие исследования структуры гена, связанные с расшифровкой генетического кода и экспериментами по изучению мутаций у различных организмов привели к пониманию того, что ген не является неделимой единицей, и мутации могут происходить и внутри гена. Больший вклад в понимание структуры гена внесли работы Маршалла У. Ниренберга по расшифровке генетического кода и его функционирования в синтезе белков, Северо Очоа, изучавшего механизмы биологического синтеза рибонуклеиновой и дезоксирибонуклеиновой кислот, а также эксперименты Сеймура Бензера и Френсиса Крика, которые в 1961 г. установили, что генетический код триплетен. Схема проведенного эксперимента следующая - проводили добавление или удаление одного нуклеотида в известную последовательность: " +" мутация - вставка одного нуклеотида. " -" мутация - выпадение одного нуклеотида. В ходе эксперимента выяснили, что: Одиночная " +" или " -" мутация в начале гена портит весь ген. Двойная " +" или " -" мутация тоже портит весь ген. Тройная " +" или " -" мутация в начале гена портит лишь его часть. Четверная " +" или " -" мутация опять портит весь ген. Этот эксперимент доказал, что код триплетен и внутри гена нет знаков препинания. Эксперимент был проведен на двух рядом расположенных фаговых генах и показал, кроме того, сто между соседними генами присутствуют знаки препинания. Дальнейшее изучение структуры генов привело к формированию понятия «генетический код» и определению общих свойств генетического кода. Под кодом ДНК, или генетическим кодом понимается отражение структуры белков в виде триплетов ДНК. Благодаря генетическому коду устанавливается однозначное соответствие между нуклеотидными последовательностями нуклеиновых кислот и аминокислотами, входящими в состав белков. Генетический код обладает следующими основными свойствами: 1.Генетический код триплетен: каждая аминокислота кодируется триплетом нуклеотидов ДНК и соответствующим триплетом иРНК. При этом кодоны ничем не отделены друг от друга (отсутствуют «запятые»). 2.Генетический код является избыточным (вырожденным): почти все аминокислоты могут кодироваться разными кодонами. Только двум аминокислотам соответствует по одному кодону: метионину (АУГ) и триптофану (УГГ). Зато лейцину, серину и аргинину соответствует по 6 разных кодонов. 3.Генетический код является неперекрывающимся: каждая пара нуклеотидов принадлежит только одному кодону (исключения обнаружены у вирусов). 4. Генетический код един для подавляющего большинства биологических систем. Однако имеются и исключения, например, у инфузорий и в митохондриях разных организмов. Поэтому генетический код называют квазиуниверсальным. 5. Компактность, или отсутствие внутригенных знаков препинания. Внутри гена каждый нуклеотид входит в состав значащего кодона.
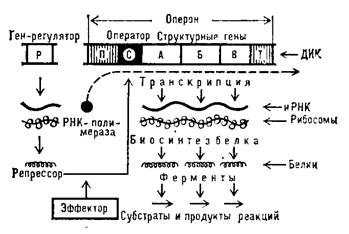
В классической генетике ген определяется не только как физическая последовательность нуклеотидов, но и как единица генетического материала, обладающая определенными свойствами: функции, мутации, рекомбинации. За время исследований, полученные данные привели к изменению первоначальных представлений о строении и свойствах генов. Изначально предполагалось, что ген - это функциональная единица, то есть нечто, определяющее отдельный признак. Такое представление сохранилось и до сих пор, но сейчас уже известно, что на один и тот же признак могут воздействовать различные гены и что при мутации гены могут давать один и тот же фенотип. Кроме того, ген определяли как единицу мутации. Эксперименты Бензера показали, что так, как ген представляет собой линейную последовательность многих участков, то мутации могут затрагивать каждый их этих составляющих. Гены изначально определялись и как единицы рекомбинаций, хотя сейчас известно, что гены не представляют собой неделимые «бусины» на цепи, а рекомбинации происходят и внутри генов. Это и следовало ожидать, если предположить, что ген представляет собой всего лишь участок ДНК, любые нуклеотидные пары которой могут изменяться, в результате мутации и рекомбинаций. Структура и регуляция генов прокариот Определение тонкой структуры гена, т.е. его организации, а также принципы работы, т.е. регуляции активности (включение-выключение), первоначально были установлены для прокариотических клеток. Эти работы выполнили Франсуа Жакоб и Жак Моно (1961; Нобелевская премия 1965). Согласно концепции Жакоба–Моно, единицей регуляции активности генов у прокариот является оперон. Оперон - функциональная единица генома у прокариот, в состав которой входят цистроны (единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами, т.е. размеры оперона превышают размеры кодирующих последовательностей ДНК. Такая функциональная организация позволяет эффективнее регулировать экспрессию (проявление) этих генов. В целом структура оперона включает: промотор, оператор, структурные гены, терминатор (рис.1).
П - Промотор – это регуляторный участок ДНК, который служит для присоединения РНК-полимеразы к молекуле ДНК. С- Оператор – это регуляторный участок ДНК, который способен присоединять белок-репрессор, который кодируется соответствующим геном. Если репрессор присоединен к оператору, то РНК-полимераза не может двигаться вдоль молекулы ДНК и синтезировать мРНК. Т- Терминатор – это регуляторный участок ДНК, который служит для отсоединения РНК-полимеразы после окончания синтеза мРНК. Транскрипция группы структурных генов, регулируется двумя элементами – геном-регулятором и оператором. Оператор часто локализуется между промотором и структурными генами; ген-регулятор может локализоваться рядом с опероном или на некотором расстоянии от него. Если продуктом гена-регулятора является белок-репрессор, его присоединение к оператору блокирует транскрипцию структурных генов, препятствуя присоединению РНК-полимеразы к специфичному участку – промотору, необходимому для инициации транскрипции. Напротив, если белком-регулятором служит активный апоиндуктор, его присоединение к оператору создает условия для инициации транскрипции. В регуляции работы оперонов участвуют также низкомолекулярные вещества – эффекторы, выступающие как индукторы либо корепрессоры структурных генов, входящих в состав оперонов. Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов). Объединение функционально близких генов в опероны, видимо, постепенно сложилось в эволюции бактерий по той причине, что у них перенос генетической информации обычно осуществляется небольшими порциями (например, при трансдукции или посредством плазмид). Значение имеет само по себе сцепление функционально родственных генов, что позволяет бактериям приобретать необходимую функцию в один этап. Структура генов эукариот Позже, когда исследователи начали изучать гены различных белков в клетках эукариот, обнаружилось, что взаимодействие генов и белков в этих организмах более сложное, чем взаимодействие генов и белков прокариот. Первые примеры такого взаимодействия были получены в 1977 году в лабораториях Филиппа Шарпа и Пьера Шамбона. Вместе со своими коллегами они гибридизировали мРНК различных генов с теми ДНК, с которых были сняты эти информационные копии. У бактерий последовательность мРНК идентична последовательности кодирующей цепи ДНК (за исключением того, что место тимина занимает урацил), поэтому структура гибридных молекул была достаточно проста. Но когда под электронным микроскопом были сделаны снимки гибридных молекул генов эукариот, то в них обнаружился ряд петель. Это значит, что мРНК и ДНК имеют не совсем идентичную последовательность, и петли были как раз теми местами, в которых они не могли соединяться. Когда последовательность мРНК сравнили с последовательностью ДНК, стало понятно, что кодирующая последовательность генов в некоторых местах прерывается некодирующей последовательностью, то есть некоторые нуклеотиды не кодируют синтез белка. Впоследствии выяснилось, что это типичная картина для ДНК эукариот. Кодирующая последовательность гена называется экзоном, а некодирующая последовательность — интроном. Некоторые гены имеют в своей структуре несколько интронов. Часто обнаруживают и такие гены, в которых больше интронов, чем экзонов.
Рис. 2. Прерывистые гены экспрессируются в виде РНК- предшественника, из которой затем удаляются интроны; экзоны, соединяясь вместе, образуют мРНК. В общем случае при транскрипции генов эукариот образуются большие молекулы РНК, содержащие как экзоны, так и интроны. Таким образом, полученные транскрипты, являются гетерогенными, т.е. они не на всем протяжении способны к трансляции. Поэтому первичные транскриптоны эукариот называются гетерогенной ядерной РНК (гяРНК) или про-мРНК. В дальнейшем происходит преобразование про-мРНК, или ее «созревание», при котором она еще в ядре вовлекается в процессинг, состоящий в том, чтоособые комплексы ферментов (сплайсингсомы) вырезают из транскрипта все интроны. Затем экзоны соединяют в одну мРНК, кодирующую производство белка. Процесс соединения экзонов называется сплайсингом (сращивание). В результате процессинга образуется «зрелая» мРНК, которая по размерам значительно короче, чем про-РНК. Молекулы про-мРНК содержат, в среднем, более 50 000 нуклеотидов, а созревшая мРНК – 500-3000 нуклеотидов. Далее полученная м-РНК транслируется как обычно. Сплайсинг характерен для эукариот и архебактерий, но иногда встречается и у прокариот. |
Последнее изменение этой страницы: 2017-05-05; Просмотров: 60; Нарушение авторского права страницы