|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Организм человека как система управленияСтр 1 из 6Следующая ⇒
Организм человека как система управления
Регуляция — управление.
Организм человека — система управления = кибернетическая система.[a]
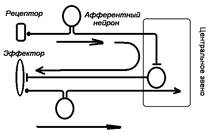
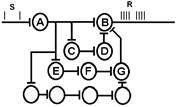
Общий план строения системы управления [1]:
В центре нашего внимания будут регуляторные (управляющие) системы функциональных систем гомеостатического уровня организации.[b]
Механизмы управления [c]
Гуморальный Нервный Нейрогуморальный
Сравнение гуморальной и нервной регуляции
Что, прежде всего, характеризует связь?
1. Скорость 2. Надёжность 3. Адресность
Гуморальная регуляция по всем этим характеристикам уступает нервной регуляции. Для гуморального механизма управления характерны относительно медленное распространение и диффузный характер управляющих воздействий, низкая надежность осуществления связи.[2] Но всегда ли нужны для регуляции функции, распространения информации такие характеристики связи?
Нервные управляющие воздействия напоминают телефонную связь, гуморальные — средства массовой информации — радио, телевиденье, прессу.
º, ¾ ¹ ', È
Разве телефоны заменили телевидение? Нет. По схожим причинам и нервная регуляция не отменяет гуморальную регуляцию.
Основные связи между нервной и гуморальной (эндокринной) системами регуляции
Осуществляются посредством взаимодействия гипоталамуса и гипофиза. [3]
Морфо‑ функциональные элементы нервной системы [4] Структурно-функциональной единицей нервной системы является нервная клетка — нейрон.
Функциональной единицей нервной системы является — рефлекторная дуга. Структурно-функциональной единицей периферической нервной системы является нервный ствол,
Спинного мозга – спинномозговой сегмент, Коры головного мозга – модуль[5], ансамбль[6], колонка. [7]
Трофическая функция тела нейрона
Перерезка аксона или дендрита ведет к гибели отростков, лежащих дистальней перерезки, а, следовательно, и синапсов этих отростков. Сома обеспечивает также рост дендритов и аксона.
Типы нейронов
Число клеточных типов неизвестно (не менее 100). Это множество, по числу элементов превышающее даже иммунную систему [13]. Классификации нейронов многочисленны. Приведем некоторые из них.
По количеству отростков
Всегда один аксон! Если отростков больше одного.
аполярные, униполярные (истинные и псевдоуниполярные, биполярные мультиполярные. [d]
Строение нейронов в значительной мере соответствует их функциональному назначению.
Истинно униполярные нейроны находятся только в мезэнцефалическом ядре тройничного нерва. Эти нейроны обеспечивают проприоцептивную чувствительность жевательных мышц. Псевдоуниполярные нейроны, на самом деле имеют два отростка (один идет с периферии от рецепторов, другой — в структуры центральной нервной системы). Оба отростка сливаются вблизи тела клетки в единый отросток. Все эти клетки располагаются в сенсорных узлах: спинальных, тройничном и т.д. Они обеспечивают восприятие болевой, температурной, тактильной, проприоцептивной, бароцептивной, вибрационной сигнализации. Биполярные нейроны имеют один аксон и один дендрит. Встречаются в основном в периферических частях зрительной, слуховой и обонятельной систем. Биполярные нейроны дендритом связаны с рецептором, аксоном — с нейроном следующего уровня организации соответствующей сенсорной системы.
Мультиполярные нейроны имеют несколько дендритов и один аксон. В настоящее время насчитывают до 60 различных вариантов строения мультиполярных нейронов, однако все они представляют разновидности веретенообразных, звездчатых, корзинчатых и пирамидных клеток.
Классификация по функциональному признаку (позиции в нейронной цепочке) [14] чувствительные (афферентные), интернейроны (вставочные), двигательные (эфферентные). Интернейроны не всегда центральные, т.е. в ЦНС!
Нейрон, по крайней мере в метасимпатической части АНС может быть одновременно вставочным и эффекторным:
Разделение на афферентные и эфферентные нейроны обозначают ещё как разделение по направлению возбуждения, соответственно к центру и от центра (периферии).
XVII век 1637 — Р.Декарт (R.Descartes, Франция) дал первое описание двигательного рефлекса. [17]
XVIII век 1784 — И.Прохаска (J.Prochaska, Чехия) развил представление о рефлексе и предложил этот термин. Описал рефлекторную дугу. [18]
XIX век 1832 — М.Голл и И.Мюллер (М.Goll, J.Muller, Германия) изложили учение о рефлексе. [19] 1863 — И.М.Сеченов (Россия) в работе «Рефлексы головного мозга» описал явление центрального торможения и распространил принцип рефлекторной реакции на психическую деятельность и поведение человека. [20]
XX век 1902 — в докладе из лаборатории И.П.Павлова впервые заявлено об открытии условных рефлексов. [21] 1941 — Н.А.Бернштейн (СССР) в трудах по физиологии движений создал схему рефлекторного кольца. [22]
Узловым моментом развития рефлекторной теории следует считать классический труд И. М. Сеченова (1863[V.G.3] ) «Рефлексы головного мозга», в котором впервые был провозглашен тезис о том, что все виды сознательной и бессознательной жизни человека представляют собой рефлекторные реакции. Рефлекс как универсальная форма взаимодействия организма и среды есть реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием центральной нервной системы.
Принципы рефлекторной теории (детерминизм [23], анализ и синтез, единство структуры и функции). Рефлекс – латинский reflexus повёрнутый, отражённый [24] Рефлекторный принцип (син. Рефлекторный механизм) [25]: 1. функционирование организма при участии рефлексов 2. основные положения рефлекторной теории 2.1. исключительное значение стимула 2.2. поступательный ход возбуждения по рефлекторной дуге (ЦНС? ) 2.3. окончание рефлекса действием.
Рефлекторное кольцо
Функциональная система
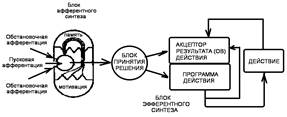
Рис. 3.1. Общая архитектура функциональной системы, определяющей целенаправленную деятельность организма на основе внутренней потребности. I — афферентный синтез; 1 — обстановочная аффсрентация, 2 — пусковая афферентация, 3 — мотивация, 4 — память; II — принятие решения; III — акцептор результатов действия; IV — программа действия (эфферентный синтез): 5 — поведенческая деятельность; V — результат поведения; VI — параметры результата; 6, 9 — обратная аффсрентация; VII — метаболизм; VIII — показатель гомеостаза; 7 — гуморальные влияния; IX — рецепторы; 8 — нервные влияния. Сплошной линией сверху обозначены границы поведенческого акта.
Свойства нервных центров Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга. Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований. Общие свойства нервных центров они перекликаются с особенностями (принципами) распространения возбуждения в ЦНС [++491, 74+].
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры, процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Суммация возбуждения и торможения.
В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана.
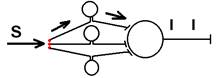
2.1.На нейроне, в области его аксонного холмика, происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона. Если с определенным интервалом к нейрону в точку А приходят импульсы, они вызывают генерацию в этой области ВПСП. Если эти ВПСП не достигают критического уровня деполяризации, то потенциал действия не возникает. 2.2.Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Приходящие в точку В, А, С нейрона (даже если они сами по себе - подпороговые), при одновременном появлении у данного нейрона могут привести к его возбуждению при условии, что суммированный ВПСП достигает или превышает критический уровень деполяризации. 2.3.Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране. Если частота следования достаточно большая, то происходит в этом месте суммация ВПСП, при достижении ВПСП критического уровня деполяризации возникает ПД, нейрон возбуждается. Это явление носит название временной суммации (происходит суммация следов возбуждения во времени). ИРРАДИАЦИЯ ВОЗБУЖДЕНИЯ От лат. Iradio, irradiare – озарять, освещать. Это распространение возбуждения из одного участка ЦНС (нервного центра) к другому. Особенности структурной организации центральных нейронов, огромное число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения. В основе иррадиации лежит дивергенция (лат. Divergentia) возбуждения. Divergentia от Divergo Divergere – направляться в разные стороны. Дивергенция – способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нейронами.[28] Если бы этот механизм не ограничивался торможением, то в целом благодаря дивергенции отсутствовала бы возможность точной координации работы ЦНС. Но тормозные процессы ограничивают дивергенцию и делают процессы управления точными. Когда торможение снимается, то имеет место полная дискоординация в деятельности ЦНС (например, при столбняке).
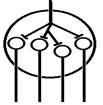
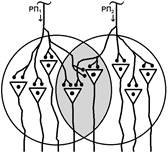
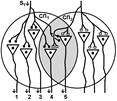
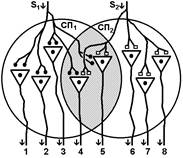
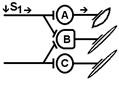
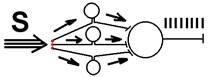
Рис. Ганглий автономной нервной системы (вегетативный ганглий[V.G.4] ) [e] Иррадиация возбуждения возможна и путем действия синаптического медиатора не только по анатомическому адресу («точка в точку», на «собственную» постсинаптическую мембрану), но и по химическому адресу при действии медиатора на рецепторы «не своих» нейронов [f]. КОНЦЕНТРАЦИИ ВОЗБУЖДЕНИЯ[V.G.5] Многие нейроны оказывают свое воздействие на один и тот же нейрон (имеет место схождение потоков импульсов к одному и тому же нейрону). Шеррингтон называл это " принцип общего конечного пути ". Например, сокращение мышцы (за счет возбуждения α ‑ мотонейрона) можно вызвать путем растяжения этой мышцы (рефлекс мышечных веретен), путем раздражения кожных рецепторов (сгибательный рефлекс) и т.п. Ч.Шеррингтон определяет физиологический механизм принципа «общего конечного пути» на уровне эфферентного звена рефлекторной дуги. В основе концентрации лежит конвергенция (лат. Convergentio) возбуждения. Convergentio от Convergo Convergere – сближать, сходиться. Конвергенция – способность двух или нескольких нейронов устанавливать синаптические связи с одним и тем же нейроном. [g] Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10: 1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные и разновалентные стимулы (свет, звук, механические раздражения и т.д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т.е. высокий уровень интеграционных функций. Различают конвергенцию нервных импульсов мультисенсорную (говорилось выше), мультибиологическую, сенсорно-биологическую и эфферентно-афферентную. Мультибиологическая конвергенция – схождение к одному нейрону двух или нескольких возбуждений от биологических раздражителей (холод, боль, жажда, половое влечение и т.п.) Сенсорно-биологическая конвергенция – схождение к одному нейрону двух или нескольких возбуждений от сенсорных и биологических раздражителей (холод, боль, жажда, половое влечение и т.п.). Это один из механизмов обучения, условных рефлексов и афферентного синтеза функциональных систем. Эфферентно-афферентная конвергенция или афферентно-эфферентная (синоним) наблюдается когда эфферентное возбуждение отходит от нейрона и взаимодействует с афферентным возбуждением, приходящим к нейрону в этот момент. Этот тип конвергенции является одним из механизмов акцептора результата действия. Окклюзия [V.G.6] Окклюзия — взаимодействие двух импульсных потоков между собой. Впервые явление окклюзии было описано Ч.Шеррингтоном. Сущность его заключается во взаимном угнетении рефлекторных реакций, при котором суммарный результат оказывается значительно меньше, чем сумма взаимодействующих реакций Окклюзия (occlusio; лат. occludo, occlusum закрывать, замыкать) Синоним – заклинивание.
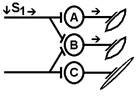
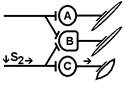
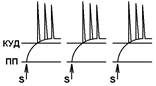
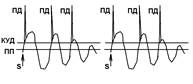
При последовательном раздражении S1 + S2 = 2 +2 = 4
При одновременном раздражении S’1 + S’2 = 3
S1 + S2 > S’1 + S’2
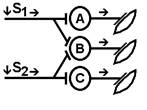
Согласно Ч.Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями взаимодействующих рефлексов. В связи с этим при одновременном поступлении двух афферентных посылок возбуждающий постсинаптический потенциал вызывается каждым из них отчасти в одних и тех же мотонейронах спинного мозга. Окклюзию используют в электрофизиологических экспериментах для определения общего звена для двух путей распространения импульсов. Если имеется общее звено, то одновременное раздражение двух путей с максимальной интенсивностью вызывает ответ меньшей величины, чем сумма ответов, получаемых при раздельном раздражении этих путей стимулами той же интенсивности. Приведем два неточных [V.G.7] объяснения явления окклюзии. Первое. «Если два нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей — феномен окклюзии» [29]
При объяснении явления окклюзии нужно говорить о перекрытии синаптических полей нервных центров, а не рецептивных полей.
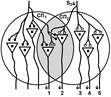
Второе. «За счет явления дивергенции один и тот же нейрон может передавать сигналы на ряд других нейронов, в результате чего возникает определенный эффект (например, происходит активация 10 мышечных волокон в мышце, в результате чего мышца развивает напряжение, равное 100 мгс. Второй нейрон возбуждает тоже 10 других волокон (100 мгс). Но если оба нейрона возбуждать одновременно, то суммарная активность мышцы будет 180 мг. Почему? Оказывается, часть волокон у них были общими (т. е. нейрон 1 и 2 передавали информацию на одни и те же волокна). Вот это явление получило название окклюзии или закупорки.» Почему это трудно признать правильным объяснением? Нейроны не конвергируют на мышце. В основе окклюзии лежит явление конвергенции. Облегчение В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т.е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолированном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций. Синоним — суммация. Да да суммация. Какая? Пространственная. [V.G.8]
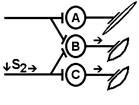
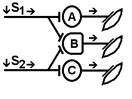
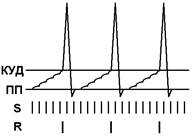
При последовательном раздражении S1 + S2 = 1 +1 = 2
При одновременном раздражении S’1 + S’2 = 3
S1 + S2 < S’1 + S’2
Последействие
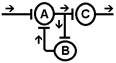
Известно, что длительность ответной реакции не соответствует времени действия раздражителя и может развиваться и осуществляться через достаточно большой промежуток времени после завершения раздражения. Один из вариантов этого свойства - длительное циркулирование импульсов по " нейронной ловушке". Итальянский физиолог Лоренто де Но обнаружил это явление: поступивший импульс может минутами или часами пробегать небольшой отрезок нейронной цепи. Благодаря этому, как полагают некоторые авторы, происходит перевод следа (энграммы) из краткосрочной памяти в долгосрочную.
Рис. Реверберация возбуждения в нервной сети (по Лоренто де Но) Предполагается, что развитие послеразрядной активности нейрона связано с особенностями проведения возбуждения через синапсы. Так как передача возбуждения в синапсах осуществляется с помощью медиаторов (в мотонейронах процесс эмиссии медиатора длится до 10 и более мс), сдвиг постсинаптического потенциала также имеет большую продолжительность. В случае развития ВПСП нейрон может разряжаться на протяжении длительного времени. Такая ритмическая импульсация затухает постепенно или обрывается внезапно. Возможен третий механизм длительного последействия, который получил название пролонгирования возбуждения. В нервных центрах существуют цепочки нейронов, связанные между собой таким образом, что импульсация определенного нейрона может вызывать генерацию импульсов в других нейронах и от них вновь приходит к первому нейрону. При этом создается возможность длительной циркуляции нервной импульсации по длинным кольцевым связям и пролонгировать возбуждение в нервном центре. Пролонгирование импульсации имеет очень большое значение в процессах переработки информации и особенно в фиксировании следов информации, т. е. памяти.
ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ (лат. transformatio преобразование, превращение)
Синоним понятия — трансформация частоты следования импульсов.
Определение понятия: Трансформация ритма возбуждений — одно из свойств проведения возбуждения в нервной системе (нервном центре), заключающееся в способности нейрона изменять ритм приходящих импульсов.[h] Существует мнение, что способность трансформировать ритм возбуждения есть только у нейрона, причём у сомы нейрона. А, например, у скелетного миоцита эта способность отсутствует. При этом следует помнить, что скелетный миоцит, как и другие эффекторные клетки, могут играть роль частотного фильтра, т.е воспринимать частоты возбуждения до какой-то предельной. Например, поступает импульс, идущий с частотой 25 Гц, а нейрон в ответ на это, возбуждаясь, генерирует 50 Гц, или наоборот, поступает 100 Гц, а выходят 40 Гц. Особенно четко проявляется трансформация ритма возбуждения при раздражении афферентного волокна одиночными импульсами. На такой импульс нейрон отвечает пачкой импульсов. [i] Выделяют несколько вероятных механизмов трансформации ритма возбуждения. [j] В ряде случаев трансформация ритма возбуждения обусловлена возникновением длительного возбуждающего постсинаптического потенциала, на фоне которого развивается несколько спайков. [k]
Этот механизм трансформации ритма возбуждения возникновением длительного возбуждающего постсинаптического потенциала, объясняет принцип кодирования «сила стимула кодируется частотой импульсов, а не амплитудой»
Другим механизмом[l] возникновения множественного разряда импульсов являются следовые колебания мембранного потенциала. Когда его величина достаточно велика, следовые колебания могут привести к достижению критического уровня деполяризации мембраны и обусловливают появление вторичных спайков. [m]
Как уже отмечалось выше способность трансформировать ритм возбудения приписывают только соме и отказывают в наличии этого свойства аксону. Аксону да, но не аксонному холмику. От состояния аксонного холмика – порога раздражения – в значительной мере может зависеть направление (урежение или учащение) и выраженность трансформации частоты следования импульсов.
Временная суммация
!!! Говоря о трансформации ритма возбуждения лучше всё же говорить не об отдельной клетке (нейроне) или отдельной её части, а о модуле (колонке для коры), ансамбле нейронов, нерном центре.
Каждый модуль, или нейронный ансамбль, представляет собой совокупность локальных нейронных сетей, которая обрабатывает информацию, передает ее со своего входа на выход, подвергает трансформации, определяемой общими свойствами структуры и ее внешними связями.[n] Кодирование[o]
!!! Говоря о трансформации ритма возбуждения лучше всё же говорить не об изменении частоты, а об изменении паттерна.
Принцип доминанты Был открыт А.А.Ухтомским на основании опытов проведенных в 1904-1911 году. Изучая ответы скелетной мышцы кошки на электрические раздражения коры больших полушарий, он обнаружил, что при акте дефекации ответы мышцы прекращаются. Проанализировав этот факт, Ухтомский пришел к мнению о наличии в ЦНС явления доминанты. Речь идет о том, что среди рефлекторных актов, которые могут быть выполнены в данный момент времени, имеются рефлексы, выполнение которых представляет наибольший " интерес" для организма, они в данный момент времени самые важные. Поэтому эти рефлексы реализуются, а другие - менее важные - тормозятся. А.А.Ухтомский назвал центры, участвующие в реализации доминантных рефлексов - " доминантным очагом возбуждения".
" Доминантный очаг" обладает рядом важных свойств: 1. он стойкий (его сложно затормозить), 2. интенсивность его возбуждения усиливается слабыми раздражителями: 3. этот очаг тормозит другие потенциальные доминантные очаги. 4.
Банунг (нем. Bahnung) — «проторение пути» суммационный рефлекс[p]
Инерционность доминанты обусловлена длительными следовыми процессами, механизмы которых детально освещены.[q] В естественных условиях длительное следовое возбуждение может быть обусловлено: 1. суммацией ВПСП приходящих подпороговых импульсов, 2. синаптической потенциацией (облегчением) при ритмическом раздражении пресинаптических входов 3. изменение концентрации К+ в снаптической щели, который как деполяризатор усиливает вхождение Ca++ в пресинаптическое окончание, 4. метаболическими следами, связанными с влиянием медиаторов на циклазные системы постсинаптических клеток, 5. циклическими связями в ЦНС, способными обеспечить следовую самостимуляцию центров.
Отчего же именно данный очаг возбуждения является доминантным? Это определяется состоянием организма, например, гормональным фоном. У голодного животного доминантными рефлексами являются пищевые. Развитием представления о доминанте являются работы Анохина о функциональной системе, в которой есть блок, принимающий решение. Именно на основе мотиваций и памяти (следов) происходит принятие решения (" Что делать в данный момент времени" ) с учетом, конечно, результатов афферентного синтеза. В настоящее время идет интенсивное изучение конкретных процессов, лежащих в основе становления и формирования доминантных очагов в ЦНС. Доминанта как один из основных принципов координационной деятельности ЦНС имеет важное значение в жизни человека. Например, именно благодаря доминанте возможно сосредоточение психической (внимание) и выполнение умственной или физической деятельности (в данном случае - это трудовая доминанта). В период поиска пищи, поедания возникает пищевая доминанта. Существуют половая, оборонительная доминанта.
СИНАПТИЧЕСКАЯ ЗАДЕРЖКА Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. [r] При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка).[s] Синаптическая задержка — замедление скорости распространения возбуждения в синапсе (межклеточном контакте) относительно скорости распространеия возбуждения в клетках возбудимых тканей (нервных, мышечных). Относительно малую скорость распространения возбуждения в синапсе связывают с длительностью процессов выделения медиатора из пресинаптического окончания, диффузии его по синаптической щели и процесса взаимодействия его с постсинаптической мембраной. [t] В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. [u] Скорость этих процессов в 10 и более раз меньше, чем скорость распространения возбуждения по нерву. С. з. в химических синапсах обычно равна 0, 2—0, 5 мс. Если учесть, что в реальных рефлекторных дугах имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд. [v]
Тонус нервного центра (фоновой активность)
Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражении определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
Пластичность
Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами. Для врача особенно важно знание этого свойства (принципа): при повреждении отдельных центров мозга их функция может перейти к другим структурам мозга (конечно, если повреждение центра не связано с наступлением смерти, что, например, бывает при нарушении дыхательного центра). Замещение утраченной функции - важнейшее приобретение ЦНС (известно, что нейроны ЦНС, как правило, не восстанавливаются) - позволяет восстанавливать утраченные свойства. Показано, что процесс возмещения утраченных функций осуществляется при обязательном участии коры больших полушарий. Показано, что у животных, которым после восстановления утраченных функций удаляли кору, вновь имела место утрата этой функции.
Рис. Спраутинг
Рис. Частичная денервация > сенсибилизация 3. Принцип субординации или соподчинения. В ЦНС имеют место иерархические взаимоотношения - начальник (кора) и подчиненные (сверху вниз - базальные ганглии, средний мозг, продолговатый, спинной) и соподчинение - нижележащий отдел подчиняется указаниям вышележащего отдела.
4. Принцип обратной связи и копий эфферентаций. Это один из важнейших принципов координации: невозможно точно координировать, управлять, если отсутствует обратная связь, т.е. данные о результатах управления. Осуществляется эта связь за счет потока импульсов с рецепторов. Этот принцип широко обсуждается в физиологии ЦНС, о нем уже говорил И.М.Сеченов, много внимания ему уделил П.К.Анохин. Копия эфферентаций. Для управления важно иметь информацию о том, какие команды посылаются на периферию. Известно, что в системах, управляющих скелетными мышцами, каждый отдел, посылая сигнал управления к работающей мышце, одновременно сообщает об этом вышележащему отделу. Это вариант обратной связи. ИНТЕГРАЦИЯ В НЕРВНЫХ ЦЕНТРАХ Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
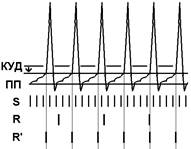
ПОСТТЕТАНИЧЕСКАЯ ПОТЕНЦИАЦИЯ Феномен посттетанической потенциации проявляется следующим образом. Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной, интенсивности. Если затем этот нерв в течение некоторого времени подвергать высокочастотному ритмическому раздражению (300—400 стимулов в секунду), то повторное редкое ритмическое раздражение приведет к резкому усилению реакции (рис. 4.4),
Рис. 4.4. Феномен посттетанической потенциации. 1 — тестовый ответ; 2 — тетаническая стимуляция; 3 — потенцированный ответ нервной клетки.
Организм человека как система управления
Регуляция — управление.
Организм человека — система управления = кибернетическая система.[a]
Общий план строения системы управления [1]:
В центре нашего внимания будут регуляторные (управляющие) системы функциональных систем гомеостатического уровня организации.[b]
Механизмы управления [c]
Гуморальный Нервный Нейрогуморальный
|
Последнее изменение этой страницы: 2019-03-30; Просмотров: 414; Нарушение авторского права страницы