|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Окисление глицерина, энергобаланс.
Глицерин в клетке подвергается фосфорилированию с участием фермента глицеролкиназы. Образующийся глицерол-3-фосфат может использоваться на анаболические реакции (синтез ТАГ, ФЛ – см. выше, а также глюкозы) и окисляться. Окисление начинается с участием НАД по a-углеродному атому. Восстановленные ДГ-азы окисляются в цепи БО, энергия БО используется в ОФ для образования АТФ. При полном окислении глицерина в чистом виде запасается 20 или 22 АТФ: 1) гликолиз – 2 АТФ; 2) окислительное декарбоксилирование ПВК – 3 АТФ; 3) АУК идет в ЦТК, энергобаланс окисления АУК – 12 АТФ; 4) в цитоплазме образуются 2 молекулы цитозольных НАДН2, которые окисляются с использованием челночных механизмов, при использовании глицерофосфатного механизма образуется 4 АТФ, при использовании малатного механизма образуется 6 АТФ. Всего образуется 21 или 23 АТФ, одна АТФ затрачивается на фосфорилирование глицерина, значит в чистом виде запасается 20 или 22 АТФ. При окислении глицерина из ФГА может образовываться ДОАФ, который кроме гликолиза используется для образования плазмологенов и тромбоцитарного активирующего фактора.
48. Регуляция метаболизма вжк ( β-окисления и биосинтеза). Синтез малонил КоА. Ацетил КоА карбоксилаза, регуляция ее активности. Транспорт ацил Ко-А через внутреннюю мембрану митохондрий. Регуляция синтеза жк. Регуляторный фермент синтеза жк – ацетил-КоА-карбоксилаза. Этот фермент регулируется несколькими способами . Активация/диссоциация комплексов субъединиц фермента. В неактивной форме ацетил-КоА-карбоксилаза представляет собой отдельные комплексы, каждый из которых состоит из 4 субъединиц. Активатор фермента-цитрат. Он стимулирует объединение комплексов, в результате чего активность фермента увеличивается . Ингибитор-пальмитоил-КоА. Он вызывает диссоциацию комплекса и снижение активности фермента. Фосфорилирование/дефосфорилирование ацетил-КоА-карбоксилазы. В постабсорбтивном состоянии или при физической работе глюкагон или адреналин через аденилатциклазную систему активируют проинкиназу А и стимулируют фосфорилирование субъединиц ацетил-КоА карбоксилазы. Фосфорилированный фермент неактивен, и синтез жирных кислот останавливается. В абсорбтивный период инсулин активирует фосфатазу, и ацетил-КоА карбоксилаза переходит в дефосфорилированное состояние. Затем под действием цитрата происходит полимеризация протомеров фермента, и он становится активным. Кроме активации фермента, цитрат выполняет и другую функцию в синтезе ЖК. В абсорбтивный период в митохондриях клеток печени накапливается цитрат, в составе которого остаток ацитила транспортируется в цитозоль. Регуляция скорости β-окисления. Β-окисление-метаболический путь , прочно связанный с работой ЦПЭ и общего пути катаболизма. Поэтому его скорость регулируется потребностью клетки в энергии, т.е. соотношениями АТФ/АДФ и NADH/NAD+ , так же, как и скорость реакции ЦПЭ и общего пути катаболизма. Скорость β-окисления в тканях зависит от доступности субстрата, т.е. от количества жирных кислот, поступающих в митохондрии. Концентрация свободных жирных кислот в крови повышается при активации липолиза в жировой ткани при голодании под действием глюкагона и при физической работе под действием адреналина. В этих условиях жирные кислоты становятся преимущественным источником энергии для мышц и печени, так как в результате β-окисления образуются NADH и ацетил-КоА, ингибирующие пируватдегидрогеназный комплекс. Превращение пирувата, образующегося из глюкозы, в ацетил-КоА замедляется. Накапливаются промежуточные метаболиты гликолиза и, в частности, глюкозо-6-фосфат. Глюкозо-6-фосфат ингибирует гексокиназу и, следовательно, препятствует использованию глюкозы в процессе гликолиза. Таким образом, преимущественное использование жк как основного источника энергии в мышечной ткани и печени сберегает глюкозу для нервной ткани и эритроцитов. Скорость β-окисления также зависит от активности фермента карнитинацилтрансферазы I. В печени этот фермент ингибируется малонил КоА, веществом , образующимся при биосинтезе жк. В абсорбтивный период в печени активируется гликолиз и увеличивается образование ацетил-КоА из пирувата. Первая реакция синтеза жк-превращение ацетил-КоА в малонил-КоА. Малонил-КоА ингибирует β-окисление жк, котрые могут использоваться для синтеза жира. Образование малонил-КоА из ацетил-КоА-регуляторная реакция в биосинтезе жк. Первая реакция синтеза жк-превращение ацетил-КоА в малонил-КоА. Фермент, катализирующий эту реакцию(ацетил Коа-карбоксилаза), относят к классу лигаз. Он содеожит ковалентно связанный биотин . В первой стадии реакции СО2 ковалентно связывается с биотином за счет энергии АТФ, во 2 стадии СОО- переносятся на ацетил-КоА с образованием малонил-КоА. Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жк. Поступивший в цитозоль цитрат активирует фермент ацетил-КоА-карбоксилазу. Малонил-КоА в свою очередь угнетает перенос высших жирных кислот из цитозоля в матрикс митохондрий, ингибируя активность внешней ацетил-КоА:карнитин-ацилтрансферазы, выключая таким образом окисление высших жирных кислот. Транспорт ацил Ко-А через внутреннюю мембрану митохондрий. Так как синтез жк происходит происходит в цитозоле клеток, то ацетил-КоА должен быть транспортирован через внутреннюю мембрану митохондрий в цитозоль. Однако внутренняя мембрана митохондрий непроницаема для ацетил-КоА, поэтому в матриксе митохондрий ацетил-КоА конденсируется с оксалоацетатом с образованием цитрата при участии цитратсинтазы : Ацетил-КоА+Оксалоацетат→ Цитрат+HS-КоА Затем транслоказа переносит цитрат в цитоплазму. Перенос цитрата в цитоплазму происходит только при увеличении количества цитрата в митохондриях, когда изоцитратдегидрогеназа и α-кетоглутаратдегидрогеназа ингибированы высокими концентрациями NADH и АТФ. Эта ситуация создается а абсорбтивном периоде, когда клетка печени получает достаточное количество источников энергии. В цитоплазме цитрат расщепляется под действием фермента цитратлиазы : Цитрат+HSКоА+АТФ→Ацетил-КоА+АДФ+Pi+Оксалоацетат Ацетил-КоА в цитоплазме служит исходным субстратом в для синтеза жк, а оксалоацетат в цитозоле подвергается превращениям, в результате которых образуется пируват.
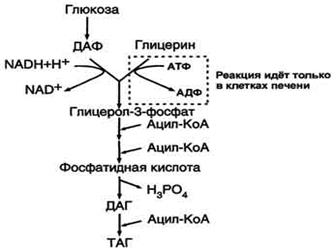
Биосинтез ТАГ (липогенез). Особенности биосинтеза ТАГ в печени и жировой ткани. Гормональная регуляция. Образование ЛПОНП в печени. Печень-основной орган, где идет синтез жирных кислот из продуктов гликолиза. В гладком Эр гепатоцитов жирные кислоты активируются и сразу же используются для синтеза жиров, взаимодействуя с глицерол-3-фосфатом. Как и в жировой ткани, синтез жиров идет через образование фосфатидной кислоты. Синтезированные в печени жиры упаковываются в ЛПОНП и секретируются в кровь.В состав ЛПОНП, кроме жиров, входят холестерол, фосфолипиды и белок апоВ-100. Это очень «длинный» белок , содержащий 11536 аминокислот. Одна молекула апоВ-100 покрывает поверхность всего липопротеина. ЛПОНП из печени секретируется в кровь, где на них, как и на ХМ, действует ЛП-липаза. Жирные кислоты поступают в ткани, в частности в адипоциты, и используются для синтеза жиров. В процессе удаления жиров из ЛПОНП под действием ЛП-липазы ЛПОНП сначала превращаются в ЛППП , а затем в ЛПНП. В ЛПНП основными липидными компонентами служат холестерол и его эфиры, поэтому ЛПНП являются липопротеинами, доставляющими холестерол в периферические ткани. Глицерол, освободившийся из липопротеинов, кровью транспортируется в печень, где опять может использоваться для синтеза жиров. Скорость синтеза ЖК и жиров в печени существенно зависит от состава пищи. Если в пище содержится белее 10% жиров, то скорость синтеза жиров в печени резко снижается. Гормональная регуляция синтеза жиров. Какой процесс будет преобладать в организме – синтез жиров или их распад зависит от поступления пищи и физической активности В абсорбтивном состоянии под действием инсулина происходит липогенез, в постабсорбтивном состоянии – липолиз, активируемый глюкагоном. Адреналин, секреция которого увеличивается при физической активности, также стимулирует липолиз. В абсорбтивный период при увеличении соотношения инсулин/глюкагон в печени активируется синтез жиров. В жировой ткани индуцируется синтез ЛП-липазы в адипоцитах и осуществляется ее экспонирование на поверхность эндотелия , следовательно в этот период увеличивается поступление жирных кислот в адипоциты. Одновременно инсулин активирует белки-переносчики глюкозы – ГЛЮТ-4. Поступление глюкозы в адипоциты и гликолиз также активируется. В результате образуются все необходимые компоненты для синтеза жиров: глицерол-3-фосфат и активные формы жирных кислот. В печени инсулин, действуя через различные механизмы, активирует ферменты путем дефосфорилирования и индуцирует их синтез. В результате увеличиваются активность и синтез ферментов, участвующих в превращении части глюкозы, поступающей с пищей в жиры. Это – регуляторные ферменты гликолиза, пируватдегидрогеназный комплекс и ферменты, участвующие в синтезе ЖК из ацетил-КоА. Результат действия инсулина на обмен углеводов и жиров в печени – увеличение синтеза жиров и секреция их в кровь в составе ЛПОНП. ЛПОНП доставляют жиры в капилляры жировой ткани, где действие ЛП-липазы обеспечивает быстрое поступление жирных кислот в адипоциты, где они депонируются в составе ТАГ.
|
Последнее изменение этой страницы: 2019-04-01; Просмотров: 562; Нарушение авторского права страницы