
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Жирные кислоты: бета-окисление в пероксисомахСтр 1 из 6Следующая ⇒
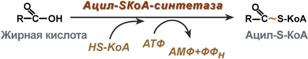
Жирные кислоты: бета-окисление в пероксисомах В пероксисомах бета-окисление жирных кислот происходит в модифицированном виде. Продуктами окисления в данном случае являются ацетил-CoA и H2O2 , последняя образуется на стадии, катализируемой связанной с флавопротеином дегидрогеназой. Этот путь окисления непосредственно не сопряжен с фосфорилированием и образованием ATP , но он обеспечивает расщепление жирных кислот с очень длинной цепью (например. C20, C22); он включается при диете, богатой жирами, или приеме гиполипидемических лекарственных препаратов , таких, какклофибрат . Ферменты пероксисом не атакуют жирные кислоты с короткими цепями, и процесс бета-окисления останавливается при образовании октаноил-CoA . Октан сильные и ацетильные группы удаляются затем из пероксисом в виде октаноилкарнитин а и ацетилкарнитин а и окисляются в митохондриях . Окисление жирных кислот (β-окисление) Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С1и С2 исходной жирной кислоты. Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом: Пальмитоил-SКоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН Этапы окисления жирных кислот 1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-S-КоА. Ацил-S-КоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЛИПИДОВ В ЖКТ. ТРАНСПОРТ ЖИРОВ ИЗ КИШЕЧНИКА ХИЛОМИКРОНАМИ. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ПИЩЕВЫХ ЛИПИДОВ С пищей в организм ежедневно поступает от 80 до 150 г липидов. Основную массу составляют жиры, наряду с глюкозой служащие главными источниками энергии. Хотя калорийность жиров значительно выше, чем углеводов (9 по сравнению с 4,7 ккал/моль), при рациональном питании жиры обеспечивают не более 30% от общего количества калорий, поступающих с пищей. Жидкие жиры (масла) содержат в своём составе полиеновые жирные кислоты, которые не синтезируются в организме; поэтому жидкие жиры должны составлять не менее одной трети жиров пищи. С липидами в организм поступают и жирорастворимые витамины A, D, Е, К. Переваривание липидов пищи происходит в кишечнике. Основные продукты гидролиза (жирные кислоты и 2-моноацилглицеролы) после всасывания подвергаются ресинтезу и последующей упаковке в хиломикроны (ХМ) в клетках слизистой оболочки кишечника. Эмульгирование жиров Жиры составляют до 90% липидов, поступающих с пищей. Переваривание жиров происходит в тонком кишечнике, однако уже в желудке небольшая часть жиров гидролизуется под действием "липазы языка". Этот фермент синтезируется железами на дорсальной поверхности языка и относительно устойчив при кислых значениях рН желудочного сока. Поэтому он действует в течение 1-2 ч на жиры пищи в желудке. Однако вклад этой липазы в переваривание жиров у взрослых людей незначителен. Основной процесс переваривания происходит в тонкой кишке. Так как жиры - нерастворимые в воде соединения, то они могут подвергаться действию ферментов, растворённых в воде только на границе раздела фаз вода/жир. Поэтому действию панкреатической липазы, гидролизующей жиры, предшествует эмульгирование жиров. Эмульгирование (смешивание жира с водой) происходит в тонком кишечнике под действием солей жёлчных кислот (рис. 8-11). Жёлчные кислоты синтезируются в печени из холестерола и секретируются в жёлчный пузырь. Содержимое жёлчного пузыря - жёлчь. Это вязкая жёлто-зелёная жидкость, содержащая главным образом жёлчные кислоты; в небольшом количестве имеются фосфолипиды и холестерол. Жёлчные кислоты представляют собой в основном конъюгированные жёлчные кислоты: таурохолевую, гликохолевую и другие (см. выше рис. 8-10). После приёма жирной пищи жёлчный пузырь сокращается и жёлчь изливается в просвет двенадцатиперстной кишки. Жёлчные кислоты действуют как детергенты, располагаясь на поверхности капель жира и снижая поверхностное натяжение. В результате крупные капли жира распадаются на множество мелких, т.е. происходит эмульгирование жира. Эмульгирование приводит к увеличению площади поверхности раздела фаз жир/вода, что ускоряет гидролиз жира панкреатической липазой. Эмульгированию способствует и перистальтика кишечника. Транспорт холестерина В плазме крови здоровых людей содержится 0,8 – 1,5 г/л ЛОНП, 3,2 – 4,5 г/л ЛНП и 1,3 – 4,2 г/л ЛВП. Липидный компонент практически всех ЛП представлен наружной оболочкой, которая образована монослоем ФЛ и холестерина и внутренним гидрофобным ядром, состоящим из ТГ и холестеридов. Кроме липидов ЛП содержат белок – аполипопротеиды А, В или С. Свободный холестерин, находящийся на поверхности ЛП, легко обменивается между частицами: меченый холестерин, введенный в плазму в составе одной группы ЛП, быстро распределяется между всеми группами. ХМ формируются в эпителиальных клетках кишечника, ЛОНП и ЛВП независимо друг от друга образуются в гепатоцитах. ЛП обмениваются своим холестерином с мембранами клеток, особенно интенсивный обмен идет между ЛП и гепатоцитами, на поверхности которых есть рецепторы для ЛПНП. Процесс переноса холестерина в гепатоциты требует энергии. Холестерин в патологии. I. Холестериноз – изменения содержания холестерина в организме. Не осложненный холестериноз – (физиологическое старение, старость, естественная смерть) проявляется накоплением холестерина в плазматических мембранах клеток в связи с уменьшением синтеза стероидных гормонов (стероидогенеза). Осложненный – атеросклероз в форме ишемической болезни сердца (инфаркт миокарда), ишемия мозга (инсульт, тромбоз), ишемия конечностей, ишемии органов и тканей, связанный с уменьшением желчегенеза. II. Изменения содержания холестерина в плазме крови. Семейная гиперхолестеринэмия – обусловлена дефектом рецепторов для ЛНП. В результате холестерин не поступает в клетки и накапливается в крови. Рецепторы по химической природе являются белками. В результате развивается ранний атеросклероз. III.Накопление холестерина в отдельных органах и тканях. Болезнь Вольмана – первичный семейный ксантоматоз – накопление эфиров холестерина и триглицеридов во всех органах и тканях, причина дефицит лизосомальной холестеринэстеразы. Ранняя смерть. Семейная гиперхолестенинэмия или βлипопротеинэмия. Нарушается поглощение ЛНП клетками, повышается концентрация ЛНП, а также холестерина. При βлипопротеинэмии наблюдается отложение холестерина в тканях, в частности в коже (ксантомы) и в стенках артерий. Отложение холестерина в стенках артерий главное биохимическое проявление атеросклероза. Вероятность заболевания атеросклерозом тем выше, чем больше отношение концентраций ЛНП и ЛВП в крови (ЛНП снабжает клетки холестерином, ЛВП удаляет из них избыток холестерина). Холестерин образует в стенках сосудов бляшки. Бляшки могут изъязвляться и язвы зарастают соединительной тканью (образуется рубец), в которую откладываются соли кальция. Стенки сосудов деформируются, становятся жесткими, нарушается моторика сосудов, суживается просвет вплоть до закупорки. Гиперхолестеринемия – главная причина отложения холестерина в артериях. Но важное значение имеют также первичные повреждения стенок сосудов. Повреждения эндотелия могут возникать в следствие гипертонии, воспалительных процессов. В области повреждения эндотелия в стенку сосудов проникают компоненты крови, в том числе липопротеиды, которые поглощаются макрофагами. Мышечные клетки сосудов начинают размножаться и тоже фагоцитировать липопротеиды. Ферменты лизосом разрушают липопротеиды, кроме холестерина. Холестерин накапливается в клетке, клетка гибнет, а холестерин оказывается в межклеточном пространстве и инкапсулируется соединительной тканью – образуется атеросклеротическая бляшка.
Методы профилактики и лечения атеросклероза направлены на уменьшение гиперхолестеринемии. Для этого применяют малохолестериновую диету, лекарства увеличивающие эксткрецию холестерина или ингибирующие его синтез, прямое удаление холестерина из крови методом гемодиффузии. Холестирамин связывает желчные кислоты и исключает их из кишечно-печеночного кровобращения, что приводит к усилению окисления холестерина в желчные кислоты. Пути биотрансформации Образование желчных кислот Синтез желчных кислот идет в эндоплазматическом ретикулуме при участии цитохрома Р450, кислорода, НАДФН и аскорбиновой кислоты. 75% холестерина, образуемого в печени, участвует в синтезе желчных кислот. Реакции синтеза желчных кислот на примере холевой кислоты
4. Этерификация холестерина. Утворення ефірів холестерину відбувається як позаклітинно, так і всередині клітини, і обидва ці процеси завжди спрямовані на те, щоб забрати молекули холестерину і транспортувати їх, або акумулювати в клітині. Позаклітинна етерифікація холестерину здійснюється ферментом лецитин-холестерин-ацилтрансферазою (ЛХАТ). Внаслідок дії ЛХАТ холестерин з поверхні ліпопротеїдів високої густини (ЛПВГ) поступає всередину ліпопротеїда у вигляді ефірів холестерину і тим самим звільняє місце для нової порції холестерину. Завдяки такому механізму ЛПВГ здатні знімати холестерин з поверхні плазматичних мембран різних клітин. Таким чином, ЛПВГ та ЛХАТ служать своєрідними уловлювачами холестерину. Біосинтез та розпад ЛХАТ та ЛПВГ відбувається в печінці, і функціональний стан печінки має значення для нормального проходження цих процесів. Клітинна етерифікація холестерину відбувається в реакції, що каталізується ацил-КоА-холестерин-ацилтрансферазою (АХАТ)-ферментом, локалізованим в багатьох органах та тканинах, однак найбільша кількість цього ферменту знаходиться в печінці та наднирниках. У наднирниках АХАТ сприяє накопиченню ефірів холестерину для синтезу стероїдних гормонів, а в печінці — виділенню з жовчю або для синтезу жовчних кислот [21]. В експериментах на тваринах встановлена наявність зворотного взаємозв'язку між секрецією холестерину з жовчю і етерифікацією холестерину в печінці та активністю АХАТ. При фармакологічному пригніченні активності АХАТ секреція холестерину з жовчю підсилюється, а при підвищенні активності АХАТ — знижується. З погляду фізіології такий зворотний зв'язок можна розглядати як наслідок нормального транспорту жовчних кислот. Жовчні кислоти прискорюють виведення вільного холестерину з жовчю, а втрата жовчних кислот призводить до підсилення синтезу холестерину в печінці та його перетворення на жовчні кислоти.Таким чином, обмін холестерину та ефірів холестерину тісно пов'язаний з печінкою та обміном жовчних кислот, а ці процеси мають відповідну залежність як від віку, так і від статі. Жирные кислоты: бета-окисление в пероксисомах В пероксисомах бета-окисление жирных кислот происходит в модифицированном виде. Продуктами окисления в данном случае являются ацетил-CoA и H2O2 , последняя образуется на стадии, катализируемой связанной с флавопротеином дегидрогеназой. Этот путь окисления непосредственно не сопряжен с фосфорилированием и образованием ATP , но он обеспечивает расщепление жирных кислот с очень длинной цепью (например. C20, C22); он включается при диете, богатой жирами, или приеме гиполипидемических лекарственных препаратов , таких, какклофибрат . Ферменты пероксисом не атакуют жирные кислоты с короткими цепями, и процесс бета-окисления останавливается при образовании октаноил-CoA . Октан сильные и ацетильные группы удаляются затем из пероксисом в виде октаноилкарнитин а и ацетилкарнитин а и окисляются в митохондриях . Окисление жирных кислот (β-окисление) Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С1и С2 исходной жирной кислоты. Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом: Пальмитоил-SКоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН Этапы окисления жирных кислот 1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-S-КоА. Ацил-S-КоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
|
Последнее изменение этой страницы: 2019-04-10; Просмотров: 396; Нарушение авторского права страницы
 Между отложением холестерина в артериях и липопротеидами крови происходит обмен, но при гиперхолестеринемии преобладает поток холестерина в стенки сосудов.
Между отложением холестерина в артериях и липопротеидами крови происходит обмен, но при гиперхолестеринемии преобладает поток холестерина в стенки сосудов.