
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Еволюція дихального апаратаСтр 1 из 5Следующая ⇒
ЛЕКЦІЯ 9 ФІЗІОЛОГІЯ ДИХАННЯ
План лекції 1. Суть процесу дихання. 2. Еволюція дихального апарата 3. Анатомічна будова системи органів дихання. Ср. 4. Легеневе дихання Механізм легеневого дихання\ 5. Характер кpовотоку в легенях 6. Зовнішнє дихання. 7. Переніс газів кров'ю 8. Клітинне дихання 9. Вплив змін умов навколишнього середовища на дихальну систему 10. Нервова регуляція дихання 11. Гуморальна регуляція дихання 12. Особливості дихання в птиць Захисні дихальні рефлекси
Суть процесу дихання. Дихання в широкому розумінні — це сукупність реакцій біологічного окиснення органічних енерговмісних речовин з виділенням енергії, необхідної для підтримання процесів життєдіяльності організму. Біологічне окиснення може відбуватись як за участю кисню (аеробне дихання), так і без нього (анаеробне дихання). Енергію, необхідну тваринам для життєдіяльності, поставляють реакції біологічного окислювання. Основною ознакою цих реакцій є переніс атомів водню з однієї молекули (донора водню) на іншу (акцептор водню) і переніс енергії на фосфорноефірний зв'язок. В клітинах тварин є ряд з'єднань, кожне з яких приймає атом водню (або його електрони) від попереднього з'єднання і віддає його наступному. Кінцевим акцептором водню в метаболізмі більшості тварин служить кисень, що утворює воду: 2Н+ + 2е + 0, 5О2 ® Н2О Організм може зберігати лише невеликий запас кисню у вигляді оксигемоглобіну крові або аналогічного йому оксиміоглобіну м'язів; тому для нормального обміну речовин організм повинен безупинно одержувати кисень. У ряді реакцій від молекул субстрату відщеплюється СО2. Цей процес, називається декарбоксилируванням, може відбуватися незалежно від використання кисню, але в більшості випадків використання кисню і відщіплення вуглекислого газу являють собою сполучені процеси. Вугільна кислота, що утворюється, повинна виводитися з клітин і рідин організму, тому що вона реагує з водою, створюючи вугільну кислоту Н2СО3. Сукупність процесів, за допомогою яких тваринні і рослинні клітини споживають кисень, віддають вуглекислоту і перекладають енергію у форму, доступну для використання, наприклад у форму енергії АТФ, називається диханням. Для дихання всім клітинам необхідне безупинне постачання киснем і видалення вуглекислоти і води. З огляду на це, можна дати таке визначення дихання: Дихання - це безупинний газообмін між організмом і зовнішнім середовищем. Дихання вищих безхребетних і хребетних — це сукупність процесів надходження кисню в організм і його споживання клітинами, а також утворення і виведення з організму вуглекислого газу. У здійсненні надходження газів беруть участь два різних механізми. Перший з них — дифузія — пасивний процес переходу дихальних газів крізь біологічні мембрани за градієнтом концентрацій — відбувається на межі між зовнішнім середовищем і кров'ю чи гемолімфою. Цей самий процес відбувається у тканинах: кисень з крові дифундує у тканинну рідину і далі до клітин, а вуглекислий газ — із клітин через тканинну рідину у кров. Другий механізм — конвекція — активне перенесення кисню і вуглекислого газу — полягає у підведенні до дихальних органів середовища (води чи повітря), що містить кисень, у перенесенні його кров'ю, гемолімфою чи целомічною рідиною від дихальних органів до тканин, а також СО2 — у зворотному напрямку. Активний транспорт газів характерний для багатоклітинних організмів. У процесі дихання більшості багатоклітинних тварин виділяють три послідовних етапи: 1) зовнішнє дихання; 2) транспорт газів кров'ю; 3) внутрішнє, або тканинне, дихання. Зовнішнє дихання, що є процесом надходження кисню із зовнішнього середовища до організму — у кров, гемолімфу або безпосередньо в тканинну чи целомічну рідину — і виведення з організму вуглекислого газу, характерне для всіх аеробних організмів. У одноклітинних організмів і нижчих багатоклітинних газообмін із зовнішнім середовищем здійснюється шляхом прямої дифузії газів через поверхню тіла. Спеціалізовані дихальні органи з'являються на досить пізніх етапах розвитку тваринного світу. До речі, дихальні рухи виникають раніше, ніж спеціалізовані дихальні органи. У тварин із шкірним диханням, наприклад у багатьох кільчастих червів, у яких немає органів дихання, спостерігаються коливні рухи тіла, не пов'язані з локомоцією, їх функція полягає у переміщенні, оновленні водного середовища навколо тіла тварини.
Еволюція дихального апарата У процесі філогенезу виробилися такі типи дихання: ¨ Дифузне, ¨ Шкірне, ¨ Кишкове, ¨ Зяброве, ¨ Трахейне, ¨ Легеневе. Коротко зупинимося на цих типах. схемой 1. Дифузне – характерно для низькоорганізованих тварин із зниженням рівнем обміну. Не потребує для свого здійснення ніяких спеціальних органів, але може здійснюватися тільки у вологому середовищі і накладає обмеження на розмір тварини. Дифузія ефективна тільки на відстані в 0, 5 - 1 мм. Цей тип дихання характерний для найпростіших – амеб, інфузорій, а також для кишковопорожнинних і плоских хробаків. 2. Шкірний тип дихання характерний для тварин декілька більшого розміру, для його здійснення необхідна хоча б примітивна кровоносна система. Шкірне дихання — це єдиний шлях газообміну у багатьох безхребетних: губок, кишковопорожнинних, більшості червів, та найпримітивніших представників різних тинів тварин. Кисень, що дифундує крізь шкіру у кільчастих червів, далі через кров надходить до всіх органів тіла. Шкірне дихання відіграє істотну роль в газообміні і у багатьох тварин, що мають спеціалізовані дихальні органи: у молюсків, голкошкірих, круглоротих, риб, амфібій і навіть у деяких плазунів. Дихальні гази (кисень і вуглекислий газ) проникають крізь біологічні мембрани лише у розчиненому стані. Якщо для водних організмів лімітуючим чинником дихання є кількість розчиненого у воді кисню, то для наземних тварин газообмінна поверхня тіла обов'язково має бути зволожена секретом шкірних залоз чи періодичним зануренням у воду (дощові черви, земноводні) 3. Кишкове дихання не має самостійного значення і виступає як доповнення до шкірного і зябрового. 4. Зябровий тип дихання характерний для багатьох тварин, що живуть у воді, зокрема для риб. При цьому типі дихання газообмін здійснюється зябрами, що подають собою численні ніжні листочки, багаті кровоносними судинами. Кисень, розчинений у воді, проникає через тонку поверхню зябер у їхні капіляри, а вуглекислий газ через зябра виділяється з крові у воду. Всі перераховані вище типи подиху характерні для водяного або вологого середовища існування. З переходом до наземного способу життя дихання починає здійснюватися з використанням трахей і легень. Чому це відбувається? Для дихання необхідна тільки волога поверхня. У умовах суші тваринам же припадає боротися з утратами рідинами. Шкірні покрови стають усе більш щільними, дифузія газів через них утруднена. Для порівняння: жаба може одержувати через шкіру біля третини необхідного їй кисню, а СО2 виводиться через шкіру більш ніж на 60 %. У людини і сільськогосподарських тварин через шкіру виводиться біля 1, 4 % СО2. Тому у тварин, що живуть на суші, в основному використовуються тільки такі органи дихання, як трахеї і легені. Трахеї – це трубочки, по котрим повітря проникає в тканини тварини, аж до окремих клітин. Така система високоефективна, але накладає істотні обмеження на розміри тварин. До трахейнодихаючих тварин відносяться комахи (бджоли, тутовий шовкопряд). 5. Трахейне дихання здійснюється спеціалізованою дихальною системою, яка доставляє кисень повітря до кожного органа, тканини, клітини без участі крові чи гемолімфи та їхніх кров'яних пігментів. Трахейна система є у комах, багатоніжок та багатьох хеліцерових 6. Легеневе дихання. У птиць і ссавців, до яких відносяться переважна більшість сільськогосподарських тварин, органами дихання є легені.
Легеневе дихання Дихання відбувається в декілька етапів. 1. Обмін повітрям між зовнішнім середовищем і альвеолами легень - атмосферне повітря при вдиху надходить в альвеоли, а частина альвеолярного повітря при видиху виділяється в зовнішнє середовище. 2. Власне легеневе дихання - газообмін між альвеолярним повітрям і кров'ю, що протікає через стінки альвеол і капілярів легень. 3. Зв'язування кисню гемоглобіном. 4. Транспорт кисню кров'ю. 5. Газообмін між кров'ю і тканинами організму. 6. Тканинне дихання. 7. Транспорт вуглекислого газу.
Повітря на шляху до легень проходить дихальні шляхи. Вони виконують в організмі ряд функцій: 1. Аналіз складу повітря, що надходить, (за рахунок наявних у носовій порожнині нюхових клітин). 2. Зігрівання або охолодження повітря (за рахунок розташованих у стінках дихальних шляхів кровоносних судин і щодо великої площі зіткнення повітря, що протікає, із поверхнями дихальних шляхів). 3. Зволоження повітря (за рахунок виділення слизу клітинами, що вистилають дихальні шляхи і секреторного епітелію дрібних залоз). 4. Очищення повітря від пилюки (за рахунок роботи миготливого епітелію і виділення слизу). Легень досягають тільки частки розміром 4 - 6 мкм. Частота биття війок складає 10 - 20 биттів у хвилину. 5. У верхніх дихальних шляхах знаходиться багато рецепторів, подразнення яких викликає захисні рефлекси – кашель, чхання, фиркання.
Механізм легеневого дихання Рух повітря у легені та з легень у навколишнє середовище обумовлене зміною тиску у середині легень. При вдиху атмосферне повітря надходить повітряносні шляхи в легені. При видиху повітря з підвищеним змістом СО2 виводяться тим же шляхом у навколишнє середовище. З трахеї повітря потрапляє до бронхів, що дихотомічно розгалужуються приблизно 23 рази. Площа поперечного перетину дихальних шляхів у міру розгалуження зростає більш ніж у 4, 5 тис. разів. У респіраторній зоні легень два середовища – газове і рідинне – розмежоване тканинами, що утворюють шляхи, по яких рухається повітря (дихальні шляхи, альвеоли) і кров (капіляри малого кола кровообігу). Легенева паренхіма і ендотелій капілярів утворюють поверхню газообміну, або альвеоло – капілярний бар'єр. Його загальна площа досягає в коня – 500 м2, у корови – 400 м2. Для порівняння, у людини вона складає 80 м2. Зміна вмісту кисню та інших газів в легенях обумовлюється вентиляцією легень за рахунок дихальних рухів. Поширення газів йде по градієнту парціального тиску – від більших значень до менших. Для здійснення вентиляції легень у більшості тварин існують дихальні м'язи, що здійснюють акт вдиху – інспірацію, та видиху – експірацію. Вдих (інспірація). Під час спокійного вдиху грудна клітка розширюється за рахунок скорочення дихальних м'язів: основних інспіраторних — діафрагми, зовнішніх міжребрових м'язів. Під час активного глибокого вдиху підключаються додаткові інспіраторні м'язи — деякі м'язи грудей, спини, м'язові волокна діафрагми скорочуються, купол діафрагми опускається і грудна порожнина збільшується, легені розширюються. У ссавців вдих відбувається під негативним тиском, тобто у плевральній порожнині тиск є негативним, і в легені засмоктується повітря. Тиск повітря в альвеолах перед вдихом і на його початку дорівнює атмосферному, а зовні легень, у плевральній щілині під час вдиху, коли грудна клітка збільшується в об'ємі, тиск падає на 6-8 мм рт. ст. Цієї різниці тиску достатньо, щоб подолати опір еластичної тяги, легень, вони розтягуються, немовби присмоктуючись до внутрішньої поверхні грудної клітки. Об'єм легень при цьому зростає, а тиск у них стає нижчим від атмосферного, і повітря надходить у легені. Видих (експірація). Поверхня розслабленої діафрагми майже вдвічі більша від площі поперечного перерізу грудної клітки, тому під час видиху вона під тиском органів черевної порожнини випнута в грудну порожнину. Спокійний видих відбувається пасивно значною мірою під впливом гравітаційної сили: зовнішні міжреброві м'язи і діафрагма розслаблюються, грудна клітка звужується, розслаблена діафрагма випинається в грудну порожнину, об'єм якої зменшується. При активному видиху до його здійснення залучаються внутрішні міжреброві м'язи. Так само активний видих має місце і під час фізичних навантажень, коли до дихальних рухів залучаються допоміжні дихальні експіраторні м'язи: косий, прямий, поперечний м'язи живота, а також м'язи, що згинають хребет. Еластичні волокна легеневої тканини, зменшуючи об'єм легень, відтягують їх разом з вісцеральною плеврою від пристінкової. Це й створює в герметичній плевральній порожнині у стані спокійного видиху негативний тиск 3-4 мм рт. ст., тобто він на 3-4 мм рт. ст. нижчий від альвеолярного, який у цей момент дорівнює атмосферному. Значення цієї негативності тиску і є мірою еластичної тяги легень. Під час видиху об'єм грудної порожнини зменшується, що зумовлює зменшення негативності плеврального тиску і спадання легень під впливом їх еластичної тяги. Типи дихання Розрізняють грудний, черевний і змішаний типи дихання. У першому випадку дихання здійснюється головним чином за допомогою м'язів грудної клітки. У диханні черевного типу переважну роль відіграють м'язи живота (косі, поперечний і прямий) і діафрагма. Черевний тип дихання є ефективнішим, оскільки скорочення діафрагми забезпечують більші зміни об'єму грудної порожнини, ніж скорочення міжребрових м'язів. Найсприятливішим для вентиляції легень є змішаний тип дихання
Всі дихальні м'язи не пов'язані з легенями. Передача відбувається пневматично завдяки плевральній порожнині, що оточує легені. Вони збільшують внутрішній об'єм грудної клітини, внутрішня поверхня якої покрита парієтальним листком плеври. Вісцеральний листок плеври щільно спаяний із тканню легень. Між цими двома листками знаходиться капілярна щілина (плевральна порожнина), заповнена плевральною рідиною. Оскільки грудна клітина являє собою замкнутий простір, а легені сполучаються з атмосферою, то тиск у легенях у стані спокою дорівнює атмосферному. У плевральній порожнині тиск на 6 — 12, 5 мм рт. ст. нижчий. Це можна визначити, ввівши в плевральну порожнину голку, сполучену з манометром. При вдиху внутришньоплевральний тиск падає ще більше – у великих тварин до – 30 – 50 мм рт. ст. Цей негативний тиск і сприяє пасивному слідуванню легень за рухами грудної клітки. Плевральна порожнина утворюється і розвивається у тварин у процесі онтогенезу. В плода легені щільно прилягають до грудної стінки і тільки після народження, у зв'язку зі зміною положення більш швидким ростом грудної клітини в порівнянні з тканню легень, утвориться різниця між обсягом грудної клітки і легень. Негативний тиск у плевральній порожнині пояснюється еластичністю самих легень. Якщо проколоти грудну клітку і впустити зовнішнє повітря в плевральну порожнину, те легеня на стороні проколу спадає. Таке явище називається пневмотораксом. При двосторонньому пневмотораксі спадаються обидві легені, і дихання стає неможливим.
Вентиляція легень Вентиляцією називають процес оновлення газового состава альвеолярного повітря під час вдиху і видиху. Інтенсивність вентиляції визначається глибиною вдиху і частотою дихальних рухів. Частота дихальних рухів в 4 – 5 разів менше за частоту скорочень серця. Частота дихання в середньому за 1 хв
При оцінці вентиляції використовують такі показники: Хвилинний об’єм дихання (Хвоб) Хвоб = Доб * n, де Доб – дихальний об’єм, тобто об’єм повітря, яке надійшло до легень за один вдих n – кількість дихальних рухів за хвилину Хвилинний об’єм дихання у коней у спокою 60 – 70 л (5 – 6*12). При легкій роботі хвилинний об’єм у коней зростає до 150 – 200 л, при тяжкий до 400 – 500 л. Життєва ємність легенів (ЖЄЛ)
Резервний (або додатковий) об’єм вдиху – після глибокого вдиху можна вдихнути ще деяку кількість повітря Резервний (або додатковий) об’єм видиху – повітря, яке можна видихнути після спокійного видоху.
Д зал (залишковий об’єм) – повітря, яке залишається у легенях навіть після смерті (легені ніколи не спадаються (роз тяжіння альвеол внаслідок негативного тиску у грудній клітині). Залишковий об’єм не входить до складу ЖЄЛ.
ЖЄЛ = Доб + Д дод.об. вд + Д дод.об. вид Загальний об’єм легенів (ЗОЛ) = ЖЄЛ + Д зал
Глибина дихання – об’єм повітря, яке надходить до легенів при кожному вдиху. Глибина дихання оборотно пропорційна частоті дихання. Частота дихання – кількість вдихів за 1 хвилину. При вдиху біля 70% повітря, що вдихається, безпосередньо надходить до легенів, а останні 25-30% не приймають участі у газообміні, тому що повітря залишається у верхніх дихальних шляхах. Відношення повітря, що вдихається до альвеолярного повітря називається коефіцієнтом легеневої вентиляції. Повітряності шляхи не приймають безпосередню участь у газообміні. Їх називають мертвим простором або анатомічно шкідливим. На вентиляцію легень, точніше альвеолярну вентиляцію, впливають ще кілька чинників. Частина альвеол вентилюється, але з різних причин не отримує крові (не перфузується). Є також альвеоли, які перфузуються кров'ю, але не вентилюються внаслідок, наприклад, локального спазму бронхів. У таких випадках газообмін не відбувається, і тому такі альвеоли утворюють альвеолярний шкідливий (мертвий) простір . Останній разом з анатомічним шкідливим простором об'єднують під назвою дихального, або функціонального (фізіологічного), шкідливого простору . Якщо об'єм анатомічного шкідливого простору є відносно стабільним, то альвеолярний шкідливий простір може змінюватися в широких межах. Наприклад, під час фізичного навантаження або у разі гіпоксії він може зменшуватись практично до нуля, тобто ця частина функціонального шкідливого простору є своєрідним резервом на випадок необхідності збільшення газообмінної поверхні легень. Крім того, через певні анатомічні причини легені вентилюються нерівномірно: верхівки легень вентилюються гірше, ніж середні й нижні частини. Це пов'язано переважно з обмеженою рухливістю ребер верхньої частини грудної клітки. Альвеолярний газообмін — це тільки один бік справи, а другий її бік — кровообіг у капілярах легень, тобто перфузія легень. Ідеальним є випадок, коли обидва процеси — і вентиляція, і перфузія легень — кількісно узгоджуються, і тоді вентиляційно-перфузійне відношення дорівнює 1. Внаслідок різних анатомічних та функціональних причин певні ділянки легень чи групи альвеол вентилюються недостатньо або зовсім не вентилюються, а інші, навпаки, за нормальної вентиляції не перфузуються кров'ю. В усіх цих випадках вентиляційно-перфузійне відношення і газообмін в альвеолах порушені. Вентиляція легень, як і перфузія, також є нерівномірною. Верхівка легень у зв'язку з малою рухомістю верхніх ребер вентилюється недостатньо або зовсім не вентилюється. І навпаки, нижні частки легень завдяки скороченням діафрагми вентилюються найкраще. Крім анатомічних чинників підтримання вентиляційно - перфузної рівноваги в легенях функціонують також локальні регуляторні механізми. Так, кровообіг у альвеолах, які не вентилюються, зменшується внаслідок звуження артеріол у відповідь на гіпоксію в альвеолах. Отже, в організмі існує чимало механізмів, спрямованих на підтримання оптимального вентиляційно-перфузійного співвідношення в легенях і на забезпечення газообміну в альвеолах, адекватного потребам організму. Таким чином, у тварин (принаймні у ссавців) у стані спокою для газообміну використовується лише частина альвеолярної поверхні легень. Резервна поверхня використовується організмом у міру потреби. У разі фізичного навантаження чи емоційного напруження внаслідок підвищення тонусу симпатичної нервової системи судини малого кола кровообігу звужуються, тиск у легеневій артерії підвищується, що спричинює збільшення газообмінної поверхні, підвищення оксигенації крові і забезпечення потреб організму в кисні. Крім того, є ще один чинник, який підвищує надійність газообмінної функції легень. Час перебування кожного еритроцита в легеневому капілярі, тобто в непрямому контакті з альвеолярним газом, коливається в межах 0, 3-0, 7 с. Однак вже через 0, 1-0, 3 с після того, як еритроцит потрапив у капіляр, він на 90 % насичується киснем. А це означає, що навіть за вдвічі більшої, ніж у спокої, швидкості руху крові в легеневих капілярах цілком достатньо часу для повноцінного газообміну. Таким чином, кров у легенях, віддавши надлишок вуглекислого газу і насичившись киснем, надходить до тканин, де відбувається протилежний процес, також виключно за законами дифузії: артеріальна кров віддає тканинам кисень і отримує від них вуглекислий газ.
Газовий склад повітря
Повітря, що видихається – це суміш повітря, що вдихається і альвеолярного повітря. Газообмін між альвеолярним повітрям і кров'ю, протікає через стінки альвеол і капілярів легень. Величина дифузії газів між альвеолами і кров’ю визначається я чисто фізичними законами, які діють у системі газ – рідина, яка розділена напівпроникною мембраною. Кількість газу, розчиненого у крові, залежить від: - Складу рідини, - Об’єму і тиску газу над рідиною, - Температурою рідини, - Фізичними властивостями даного газу. Коефіцієнт розчинності відображає об’єм газу, який можна розчинити в 1 мл. рідини при t=0º С і тиску 760 лм. рт. ст. Якщо над рідиною знаходиться декілька газів, то кожен з них розчиняється відповідно його парціального тиску. Парціальний тиск – це частина загального тиску газової суміші, яка приходиться на долю того або іншого газу суміші (тобто це тиск газу, що входить до складу газової суміші, яке він оказував би при тій самій температурі, заміщуючи один весь об’єм). Виражається у паскалях або мм рт.ст. p=P(a: 100), де Р – загальний тиск газової суміші, мм рт.ср. а – вміст газу в об’ємних процентах (об%) р – парціальний тиск газу
при Р=760, t = 22º С р О2 = 0, 21*760=159 мм рт ст. (21, 2 кПа) р СО2 = 0, 0003*760=0, 23 мм рт ст. (0, 003 кПа) р N2 = 0, 78*760=595 мм рт ст. (0, 8 кПа)
Для рідини замість терміна “тиск” використовують “напруга газу” тому що розчинений у рідини газ не оказує тиску, який можна виміряти. Оскільки розчинений газ знаходиться у рівновазі зі своїм газовим середовищем, умовно вважається, що газ у рідині знаходиться під тим самим парціальним тиском. Парціальний тиск, мм рт ст |
Загальний тиск | |||||||||||||||||||||||||||||||||||||||
| Н2О | СО2 | О2 | N 2 | |||||||||||||||||||||||||||||||||||||
| Повітря, що вдихається | 6 | 0, 3 | 159 | 595 | 760 | |||||||||||||||||||||||||||||||||||
| Альвеолярне повітря | 47 | 40 | 100 | 573 | 760 | |||||||||||||||||||||||||||||||||||
| Повітря, що видихається | 47 | 32 | 116 | 565 | 760 | |||||||||||||||||||||||||||||||||||
| Артеріальна кров | 47 | 40 | 95 | 573 | 755 | |||||||||||||||||||||||||||||||||||
| Змішана венозна кров | 47 | 46 | 40 | 573 | 706 | |||||||||||||||||||||||||||||||||||
| Тканинна рідина | 47 | 60 | 20 | 573 | 700 | |||||||||||||||||||||||||||||||||||
Градієнт парціального тиску (різниця парціального тиску) – це основний фактор, який визначає дифузію газів.
Різниця між тиском О2 в альвеолярному повітрі та напругою його у венозній крові легенів:
100-40=60 мм рт ст.
Різниця між напругою СО2 у венозній крові та тиском СО2 в альвеолярному повітрі:
46-40=6 мм рт ст.
Але розчинність СО2 у тканинах легеневих мембран (аерогематичний бар’єр) вище розчинності О2 більш, ніж у 20 разів.
Тиск кисню у венозній крові, що надходить до легеневих капілярів, значно нижче, а тиск вуглекислоти вище, ніж альвеолярному повітрі. Ця різниця тисків забезпечує дифузію кисню з альвеолярного повітря у кров, а вуглекислоти – з крові в альвеолярне повітря.
Під час одного вдиху тиск вуглекислоти у крові та альвеолярному повітрі встигає дійти до стану рівноваги, а тиск кисню в артеріальній крові залишається нижчим, ніж в альвеолярному повітрі. Це залежить від більшої швидкості дифузії СО2 у порівнянні з киснем.
Під час роботи тиск СО2 у венозній крові зростає до 50-60 мм рт ст. У результаті капілярно – альвеолярний градієнт тиску СО2 при роботі може збільшуватися у 2-3 рази, що полегшує обмін газів у легенях, не дивлячись на значне прискорення у них кровотоку і, відповідно, зменшення часу знаходження крові в альвеолярних капілярах.
Зв’язування та переніс кров’ю кисню
Кисень надходить до крові, у незначній кількості поглинається плазмою і в основному – гемоглобіном.
Hb+O2=HbO2 (оксигемоглобін) – нестійке поєднання, що легко дисоціює
Зв’язування киснем гемоглобіну залежить від:
1) від напруги кисню у крові. При зниженні напруги кисню, оксигемоглобін віддає весь кисень при низькому тиску у тканинах
2) присутність СО2. при однаковому тиску, але у присутності СО2 зв’язується менша кількість кисню. Тому надходження СО2до крові з тканин полегшує вивільненні кисню з оксигемоглобіну та перехід його у тканини, а виділення СО2 з крові в легенях полегшує зв’язування кисню гемоглобіном.
Киснева ємність крові – це кількість кисню, яке може бути пов’язане 100 мл крові при повному переході гемоглобіну крові у оксигемоглобін. (1гр гемоглобіну пов’язує 1, 34 мл кисню.)
Зв’язування та переніс кров’ю СО2
У венозній крові вміст СО2 складає 50-58%, найбільша самотина – у плазмі і еритроцитах у вигляді вугільної кислоти; 2, 5об% у розчиненому стані і 4-5об% у вигляді HbCO2.
СО2 з тканин легко дифундує у кров, тому що напруга СО2 у тканинах вище його напруги у венозній крові. Вуглекислота, розчиняючись у плазмі, дифундує всередину еритроцитів, де під впливом карбоангідрази перетворюється у вугільну кислоту:
СО2 +Н2О ↔ Н2СО3
Тому що вся вуглекислота в еритроцитарній плазмі перетворюється на вугільну кислоту (Н2СО3) то напруга СО2 в середині еритроцитів дорівнює 0. тому в еритроцити постійно надходять нові порції вуглекислоти. Концентрація іонів НСО3, які утворюються в еритроцитах зростає, і ці іони починають дифундувати в плазму, де:
НСО3 +NaCl → NaHCO3 +Cl
Cl проникає у еритроцити.
Крім утворення HbO2 в еритроцитах відбувається віддача O2 оксигемоглобіну:
КHbO2 + Н2СО3= НHb + О2 +КНСО3
Клітинне дихання
З фізіологічного погляду тканинне дихання — це обмін дихальними газами між кров'ю і тканинами, що відбувається у процесі біологічного окиснення органічних речовин у клітинах організму. При цьому відбувається поглинання тканинами кисню і виділення вуглекислого газу, а внаслідок окиснення й розпаду поживних речовин виділяється енергія, необхідна для підтримання життя і діяльності організму. За нормальних умов існування при достатній кількості кисню майже вся енергія утворюється внаслідок аеробного процесу окислення, який постачає в 10-15 разів більше енергії, ніж анаеробний процес гліколізу.
Біологічне окиснення відбувається в мітохондріях, де містяться ферменти дихального ланцюга. Для їх нормальної функції потрібно, щоб напруга кисню (рО2) в мітохондріях була не нижчою за 1 мм рт. ст., інакше окиснення субстрату й отримання енергії для життєвих процесів припиняється.
Надходження кисню до тканин здійснюється як шляхом конвекції (перенесення кисню кров'ю, тканинною рідиною, через цитоплазму всередині клітини), так і за допомогою дифузії. Дифузія відбувається і в рідинах, що переносять кисень, проте головним чином забезпечує перехід кисню крізь біологічні мембрани: стінку капіляра, мембрани клітин та мітохондрій. Оскільки кисень добре розчиняється в ліпідах, він легко проникає крізь біологічні мембрани.
Рушійною силою дифузії газів, як і в альвеолярному газообміні, є градієнт концентрацій — напруги газів по обидва боки мембрани. В артеріальному кінці капіляра рО2 становить 90 мм рт. ст., а в тканинах — 20-40 мм рт. ст. Значення рСО2 в тканинах досягає 60 мм рт. ст., а в артеріальній крові — 40 мм рт. ст. І хоча різниця напруг для СО2 втричі менша, ніж для О., (відповідно 20 і 60 мм рт. ст. ), вона в обох випадках є цілком достатньою для переходу СО2 з тканин у кров та забезпечення тканин необхідною кількістю кисню. Процес дифузії триває до вирівнювання концентрацій між середовищами. Тому кров не може віддати весь кисень тканинам, так само тканини не можуть цілком звільнитись від СО2 — перехід цих газів з одного середовища до іншого припиняється, коли їх напруга в середовищах зрівнюється.
Кількість кисню, що надходить до органа чи тканини, залежить від їх кровопостачання, а рівень кровопостачання органів, у свою чергу, зумовлюється особливостями їх функції, інтенсивністю метаболізму, функціональним станом.
Споживання кисню тканинами. Різні органи і тканини в стані спокою організму споживають кисень з неоднаковою інтенсивністю, що пов'язано з виконуваною ними роботою. Так, найінтенсивніше споживають кисень кора великого мозку, серце і кіркова речовина (кора) нирок — органи, які працюють безперервно, навіть під час сну. Найменше потребують кисню шкіра, скелетні м'язи та внутрішня мозкова речовина нирок, їх функціональне навантаження у спокійному стані організму мінімальне.
Існує досить чітка залежність між інтенсивністю споживання кисню органами і їх кровопостачанням у стані спокою. Така залежність ще більшою мірою виявляється під час збільшення функціональної активності організму. Так, під час тяжкого фізичного навантаження людини кровопостачання серця зростає в 4-6 разів, а скелетних м'язів - у 20-30 разів і досягає значень відповідно 400 і 70 мл/хв на 100 г тканини.
З кров'ю до органів надходить кисень, але вони використовують його не повністю. Коефіцієнт утилізації кисню, що характеризує частку спожитого органом кисню від загального його вмісту в артеріальній крові, також свідчить про неоднакову інтенсивність дихання різних органів.
Коефіцієнт утилізації кисню – це кількість кисню, яке отримують тканини із загального вмісту в артеріальній крові, виражене у процентах
Кут. О2 =О2арт-О2 веноз
Підвищенню Кут сприяє посилене утворення молочної та вугільної кислот при значній фізичній роботі, а також розкриття не функціонуючих капілярів у працюючій тканині.
Так, для більшості органів у стані спокою коефіцієнт утилізації кисню становить 0, 3-0, 4, для серця — 0, 6, у напружено функціонуючих м'язах він зростає до 0, 7-0, 9, а з печінці може досягти навіть 0, 97.
Клітини органів споживають лише кисень, розчинений у тканинній рідині. Однак його там дуже мало, і тканини для нормального функціонування потребують постійного надходження кисню з кров'ю. Тільки скелетні м'язи й міокард завдяки наявності в них пігменту міоглобіну, який зв'язується з киснем, здатні резервувати певну кількість кисню.
У деяких тварин, особливо водних ссавців, міоглобіну в скелетних м'язах майже.тільки, скільки й гемоглобіну в крові, завдяки чому вони можуть довго перебувати під водою за рахунок кисню, зв'язаного міоглобіном. Проте у інших ссавців вміст міоглобіну в м'язах невеликий, і, отже, депонованого в них кисню вистачає ненадовго. Що стосується серця, то в разі повного припинення його кровопостачання весь кисень у ньому використовується через 3-4 секунди і окисні процеси припиняються.
Звичайно, такий малий резерв кисню з міокарді не врятує організм від смерті у разі тривалого припинення кровопостачання серця, проте він відіграє надзвичайно важливу роль в ті короткі моменти серцевого циклу, коли під час систоли волокна міокарда перетискають дрібні вінцеві судини і кровотік у товщі міокарда на 0, 2-0, 3 с різко знижується або зовсім припиняється.
Завдяки наявності міоглобіну і здатності міокарда використовувати енергію гліколізу серце тварини може працювати ще кілька десятків секунд після припинення кровопостачання. На відміну від серця головний мозок, позбавлений будь-яких запасів кисню і маючи високий рівень його споживання, виявляє надзвичайно високу чутливість до нестачі кисню. Навіть короткочасне перетиснення однієї з двох сонних артерій, через які кров живить мозок, призводить до миттєвої втрати свідомості.
За умов нормального функціонування органа між споживанням кисню клітинами і надходженням його встановлюється рівновага, що відображається певним рівнем напруги кисню (рО2) в органі. Його вимірюють за допомогою полярографічного методу, використовуючи для цього тонкі платинові електроди. Напруга кисню в різних точках будь-якого органа може коливатись у широких межах — від 0 до 80 мм рт. ст. У той самий час середні значення рО2 для скелетних м'язів, печінки і кори великого мозку мало відрізняються і перебувають у діапазоні 26-32 мм рт. ст., що свідчить не стільки про інтенсивність дихання, скільки про рівень постачання їх киснем.
Широкий діапазон значень рО2 в різних точках одного й того самого органа ілюструє явище, описане Крогом під назвою " гра капілярів. Високі значення рО2 реєструються в тих ділянках органа, де тканину живлять відкриті капіляри, а там, де капіляри тимчасово закриті, спостерігаються низькі або й нульові значення напруги кисню.
Регуляція тканинного дихання і постачання тканини киснем. Тканинне дихання тісно пов'язане з функціональним станом органа чи тканини. Такий зв'язок здійснюється як безпосередньо: функція — обмін речовин — дихання, так і через окремі незалежні шляхи регуляції обміну речовин і дихання в органах, наприклад активація глікогенолізу під час фізичного навантаження з подальшим окисненням глюкози під впливом симпатичної нервової системи та адреналіну надниркових залоз. В обох випадках у регуляції бере участь центральна нервова система, зокрема гіпоталамус, який здійснює регуляцію обміну речовин в організмі через автономну нервову систему і залози внутрішньої секреції. Так, подразнення деяких структур гіпоталамуса може посилити споживання кисню організмом чи окремим його органом, наприклад печінкою.
Між споживанням кисню тканинами і його надходженням до тканин існує тісний зв'язок. У переважній більшості випадків надходження кисню до органів регулюється змінами кровопостачання, які забезпечуються головним чином периферичними механізмами. Основну роль у цьому процесі відіграють метаболіти, гіпоксія, СО2, аденозин тощо. Вони розширюють судини функціонуючих органів і збільшують приплив крові до них. Проте коли периферичні місцеві механізми охоплюють великі судинні ділянки, що може загрожувати зниженням артеріального тиску, вмикаються центральні механізми регуляції. Так, для здійснення захисної реакції залучається велика кількість скелетних м'язів, і щоб розширення їхніх судин не призвело до падіння артеріального тиску, центр захисних реакцій у гіпоталамусі надсилає команди на звуження судин в органах травного каналу й нирок і одночасне посилення роботи серця. Таким чином забезпечується адекватне потребам працюючих м'язів надходження О2. Печінка займає виняткове становище щодо забезпечення її функцій киснем. Більшу частину потрібного їй кисню вона отримує через ворітну вену, кровотік якої регулюється судинами травного каналу. Оскільки артеріальний кровотік у печінці не може повністю компенсувати зменшене надходження кисню ворітною системою у разі звуження судин травного каналу, печінка задовольняє свої потреби в кисні шляхом посиленої екстракції кисню — відбору О2 з артеріальної та ворітної крові. У разі зменшення кровотоку у ворітній системі об'ємна частка О2 в крові печінкових вен знижується до 2-4 %, що свідчить про високий рівень утилізації кисню.
Регуляція дихання
У понятті регуляції дихання розрізняють два аспекти: це, по-перше, підтримка ритмічності дихання, тобто механізм автоматії дихального центру, по-друге, процеси власне регуляції дихання, тобто модуляції функції дихального центру, її пристосування до потреб організму.
Дихання – саморегулюючий процес, у якому головне значення має дихальний центр, розташований у ретикулярної формації довгастого мозку, в області дна четвертого мозкового шлуночка (Н.А. Миславский, 1885).
Дихальний центр — це сукупність нейронів центральної нервової системи, які забезпечують ритмічну зміну вдиху видихом і навпаки, а також пристосовують частоту і глибину дихання до мінливих потреб організму і відповідно до умов його існування.
На підставі дослідів зі зруйнуванням стовбура мозку чи відокремленням головного мозку від спинного ще в XVIII ст. було отримано перші відомості про дихальний центр і його локалізацію. Наприкінці XIX ст. М. О. Міславський, а згодом і В. М. Бехтерев установили, що дихальний центр розміщений на дні IV шлуночка і складається з центру вдиху ( енспіраторного ) та центру видиху ( експіраторного ). Було також установлено, що дихальний центр парний, симетричний, розміщений по обидва боки від шва довгастого мозку.
У наш час у ділянках довгастого мозку, які методом електричного подразнення ідентифікувались як інспіраторний та експіраторний центри, виявлено невелику кількість (10-15 %) нейронів, імпульси яких були синхронними з диханням. Це дихальні нейрони . Одні з них давали залп імпульсів під час вдиху — інспіраторні дихальні нейрони , серед яких розрізняють ранні та пізні інспіраторні нейрони, інші виявляли свою активність під час видиху — експіраторні нейрони. І ті, й інші досить нерівномірно локалізуються в більшій частині структур довгастого мозку, але в окремих ділянках спостерігається переважна концентрація певних дихальних нейронів.
В обох дихальних групах аксони частини нейронів спускаються низхідними шляхами до дихальних мотонейронів спинного мозку. Це бульбоспінальні нейрони. Відростки решти дихальних нейронів не виходять за межі довгастого мозку, з'єднуючись між собою і з бульбоспінальними нейронами. Бульбоспінальні нейрони обох дихальних груп — це головним чином повні інспіраторні нейрони, функція яких полягає в передачі до мотонейронів спинного мозку імпульсів з дихального центру. Щодо ранніх та пізніх інспіраторних нейронів, то вони належать до власних пропріобульбарних нейронів довгастого мозку, які, взаємодіючи між собою, забезпечують ритмічну діяльність дихального центру довгастого мозку.
рис
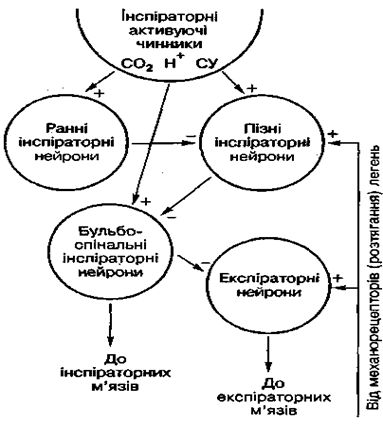
Мал. 58. Схема роботи дихального центру:
" +" — збуджувальний вплив; " -" — гальмівний вплив; СУ — сітчастий утвір
Пневмотаксичний центр.
У верхній третині моста (варолієвого) розміщені скупчення дихальних нейронів, які функціонально відрізняються від дихальних нейронів довгастого мозку і утворюють пневмотаксичний центр. Вони не жорстко пов'язані з дихальним ритмом, можуть змінювати свою функцію: з інспіраторних перетворюватись на експіраторні і навпаки. Велика кількість нейронів активізується в перехідних фазах від вдиху до видиху чи від видиху до вдиху. Електричне подразнення пневмотаксичного центру може спричинювати як вдих, так і видих. Пневмотаксичний центр отримує імпульси від інспіраторного центру, а свої сигнали надсилає до обох — інспіраторного та експіраторного центрів. Зруйнування цього центру або відокремлення його від довгастого мозку призводить до сповільнення дихання.
Крім цього, у регуляції дихання беруть участь і відділи мозку, що розташовані вище, що більш тонко пристосовують дихання до зміни зовнішніх і внутрішніх умов.
Це:
1. Середній мозок – переключення інформації з пропріо- і інтерорецепторов до дихального центру.
2. Проміжний мозок – переключення інформації про інтенсивність метаболізму.
3. Мозочок – тонізуючий вплив.
4. Кора головного мозку підкоряє дихання довільним впливам. Під впливом кори головного мозку утворяться умовні рефлекси, що регулюють дихання. (У коней перед стартом дихання частішає). По пірамідних шляхах імпульс до мотонейронам проходить, минаючи бульбарний центр.
Особливості дихання птиць
Особливості дихання птиць пов'язані з особливостями життя й обміну речовин у цих тварин і особливістю будови їхніх дихальних органів. Ці розходження визначаються способом життя птиць (пристосованістю до польоту або плавання), а також інтенсивним обміном речовин.
Захисні дихальні рефлекси
При подразненні рецепторів слизуватої оболонки порожнини носа, гортані, трахеї пилом, слизом, або іншими подразниками.
Кашель – це короткий глибокий вдих
Чихання - глибокий вдих, який змінюється різким коротким видохом.
ЛІТЕРАТУРА
Основна
1. Георгиевский В.И. Физиология сельскохозяйственных животных. – М.: Агропромиздат, 1990. – 511 с.
2. Науменко В.В. та ін. Фізіологія сільськогосподарських тварин: Підручник. – Київ: Вид-во " Сільгоспосвіта", 1994. – 512 с.
3. Фізіологія сільськогосподарських тварин: Практикум. – К.: Либідь, 1993. – 224 с.
Додаткова
1. Мелехин Г.П., Гридин Н.Я. Физиология сельскохозяйственной птицы. – М.: Колос, 1977. – 287 с
2. Тыщенко В.П. Физиология насекомых. – М.: Высш. шк., 1986. – 303 с.
3. Воронцов Д.С., Ємченко А.І. Фізіологія тварин і людини. – К.: Радянська школа, 1952. – 655 с.
4. Кіндя В.І., Куровський Ю.А., Мусієнко В.Ф. Словник-довідник з анатомії та фізіології с.-г. тварин. – К.: " Урожай", 1993. – 294 с.
5. Рихтер и др. Основные физиологические показатели у животных и технология содержания. – М.: Колос, 1982. – 192 с.
6. Стреси сільськогосподарських тварин і птиці. – К.: Урожай, 1990. – 144 с.
7. Шмидт-Ниельсен К. Физиология животных. Приспособление и среда. Книги 1, 2. – М.: Мир, 1982. – 800 с.
8. Шмидт-Ниельсен К. Размеры животных: почему они так важны? – М.: Мир, 1987. – 259 с.
9. Физиология сельскохозяйственных животных. -М.: Агропромиздат, 1991. - 432 с.
10. Общий курс физиологии человека и животных. В 2 кн. Кн. 2. Физиология висцеральных систем. - М.: Высш. шк., 1991. - 528 с.
ЛЕКЦІЯ 9
ФІЗІОЛОГІЯ ДИХАННЯ
План лекції
1. Суть процесу дихання.
2. Еволюція дихального апарата
3. Анатомічна будова системи органів дихання. Ср.
4. Легеневе дихання
Механізм легеневого дихання\
5. Характер кpовотоку в легенях
6. Зовнішнє дихання.
7. Переніс газів кров'ю
8. Клітинне дихання
9. Вплив змін умов навколишнього середовища на дихальну систему
10. Нервова регуляція дихання
11. Гуморальна регуляція дихання
12. Особливості дихання в птиць Захисні дихальні рефлекси
Суть процесу дихання.
Дихання в широкому розумінні — це сукупність реакцій біологічного окиснення органічних енерговмісних речовин з виділенням енергії, необхідної для підтримання процесів життєдіяльності організму. Біологічне окиснення може відбуватись як за участю кисню (аеробне дихання), так і без нього (анаеробне дихання).
Енергію, необхідну тваринам для життєдіяльності, поставляють реакції біологічного окислювання. Основною ознакою цих реакцій є переніс атомів водню з однієї молекули (донора водню) на іншу (акцептор водню) і переніс енергії на фосфорноефірний зв'язок. В клітинах тварин є ряд з'єднань, кожне з яких приймає атом водню (або його електрони) від попереднього з'єднання і віддає його наступному. Кінцевим акцептором водню в метаболізмі більшості тварин служить кисень, що утворює воду:
2Н+ + 2е + 0, 5О2 ® Н2О
Організм може зберігати лише невеликий запас кисню у вигляді оксигемоглобіну крові або аналогічного йому оксиміоглобіну м'язів; тому для нормального обміну речовин організм повинен безупинно одержувати кисень.
У ряді реакцій від молекул субстрату відщеплюється СО2. Цей процес, називається декарбоксилируванням, може відбуватися незалежно від використання кисню, але в більшості випадків використання кисню і відщіплення вуглекислого газу являють собою сполучені процеси. Вугільна кислота, що утворюється, повинна виводитися з клітин і рідин організму, тому що вона реагує з водою, створюючи вугільну кислоту Н2СО3. Сукупність процесів, за допомогою яких тваринні і рослинні клітини споживають кисень, віддають вуглекислоту і перекладають енергію у форму, доступну для використання, наприклад у форму енергії АТФ, називається диханням. Для дихання всім клітинам необхідне безупинне постачання киснем і видалення вуглекислоти і води. З огляду на це, можна дати таке визначення дихання: Дихання - це безупинний газообмін між організмом і зовнішнім середовищем.
Дихання вищих безхребетних і хребетних — це сукупність процесів надходження кисню в організм і його споживання клітинами, а також утворення і виведення з організму вуглекислого газу. У здійсненні надходження газів беруть участь два різних механізми.
Перший з них — дифузія — пасивний процес переходу дихальних газів крізь біологічні мембрани за градієнтом концентрацій — відбувається на межі між зовнішнім середовищем і кров'ю чи гемолімфою. Цей самий процес відбувається у тканинах: кисень з крові дифундує у тканинну рідину і далі до клітин, а вуглекислий газ — із клітин через тканинну рідину у кров.
Другий механізм — конвекція — активне перенесення кисню і вуглекислого газу — полягає у підведенні до дихальних органів середовища (води чи повітря), що містить кисень, у перенесенні його кров'ю, гемолімфою чи целомічною рідиною від дихальних органів до тканин, а також СО2 — у зворотному напрямку. Активний транспорт газів характерний для багатоклітинних організмів.
У процесі дихання більшості багатоклітинних тварин виділяють три послідовних етапи: 1) зовнішнє дихання; 2) транспорт газів кров'ю; 3) внутрішнє, або тканинне, дихання.
Зовнішнє дихання, що є процесом надходження кисню із зовнішнього середовища до організму — у кров, гемолімфу або безпосередньо в тканинну чи целомічну рідину — і виведення з організму вуглекислого газу, характерне для всіх аеробних організмів. У одноклітинних організмів і нижчих багатоклітинних газообмін із зовнішнім середовищем здійснюється шляхом прямої дифузії газів через поверхню тіла. Спеціалізовані дихальні органи з'являються на досить пізніх етапах розвитку тваринного світу. До речі, дихальні рухи виникають раніше, ніж спеціалізовані дихальні органи. У тварин із шкірним диханням, наприклад у багатьох кільчастих червів, у яких немає органів дихання, спостерігаються коливні рухи тіла, не пов'язані з локомоцією, їх функція полягає у переміщенні, оновленні водного середовища навколо тіла тварини.
Еволюція дихального апарата
У процесі філогенезу виробилися такі типи дихання:
¨ Дифузне,
¨ Шкірне,
¨ Кишкове,
¨ Зяброве,
¨ Трахейне,
¨ Легеневе.
Коротко зупинимося на цих типах. схемой
1. Дифузне – характерно для низькоорганізованих тварин із зниженням рівнем обміну. Не потребує для свого здійснення ніяких спеціальних органів, але може здійснюватися тільки у вологому середовищі і накладає обмеження на розмір тварини. Дифузія ефективна тільки на відстані в 0, 5 - 1 мм. Цей тип дихання характерний для найпростіших – амеб, інфузорій, а також для кишковопорожнинних і плоских хробаків.
2. Шкірний тип дихання характерний для тварин декілька більшого розміру, для його здійснення необхідна хоча б примітивна кровоносна система. Шкірне дихання — це єдиний шлях газообміну у багатьох безхребетних: губок, кишковопорожнинних, більшості червів, та найпримітивніших представників різних тинів тварин. Кисень, що дифундує крізь шкіру у кільчастих червів, далі через кров надходить до всіх органів тіла. Шкірне дихання відіграє істотну роль в газообміні і у багатьох тварин, що мають спеціалізовані дихальні органи: у молюсків, голкошкірих, круглоротих, риб, амфібій і навіть у деяких плазунів. Дихальні гази (кисень і вуглекислий газ) проникають крізь біологічні мембрани лише у розчиненому стані. Якщо для водних організмів лімітуючим чинником дихання є кількість розчиненого у воді кисню, то для наземних тварин газообмінна поверхня тіла обов'язково має бути зволожена секретом шкірних залоз чи періодичним зануренням у воду (дощові черви, земноводні)
3. Кишкове дихання не має самостійного значення і виступає як доповнення до шкірного і зябрового.
4. Зябровий тип дихання характерний для багатьох тварин, що живуть у воді, зокрема для риб. При цьому типі дихання газообмін здійснюється зябрами, що подають собою численні ніжні листочки, багаті кровоносними судинами. Кисень, розчинений у воді, проникає через тонку поверхню зябер у їхні капіляри, а вуглекислий газ через зябра виділяється з крові у воду. Всі перераховані вище типи подиху характерні для водяного або вологого середовища існування. З переходом до наземного способу життя дихання починає здійснюватися з використанням трахей і легень.
Чому це відбувається?
Для дихання необхідна тільки волога поверхня. У умовах суші тваринам же припадає боротися з утратами рідинами. Шкірні покрови стають усе більш щільними, дифузія газів через них утруднена. Для порівняння: жаба може одержувати через шкіру біля третини необхідного їй кисню, а СО2 виводиться через шкіру більш ніж на 60 %. У людини і сільськогосподарських тварин через шкіру виводиться біля 1, 4 % СО2.
Тому у тварин, що живуть на суші, в основному використовуються тільки такі органи дихання, як трахеї і легені.
Трахеї – це трубочки, по котрим повітря проникає в тканини тварини, аж до окремих клітин. Така система високоефективна, але накладає істотні обмеження на розміри тварин. До трахейнодихаючих тварин відносяться комахи (бджоли, тутовий шовкопряд).
5. Трахейне дихання здійснюється спеціалізованою дихальною системою, яка доставляє кисень повітря до кожного органа, тканини, клітини без участі крові чи гемолімфи та їхніх кров'яних пігментів. Трахейна система є у комах, багатоніжок та багатьох хеліцерових
6. Легеневе дихання. У птиць і ссавців, до яких відносяться переважна більшість сільськогосподарських тварин, органами дихання є легені.
Последнее изменение этой страницы: 2019-06-08; Просмотров: 110; Нарушение авторского права страницы
