|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Сколько воды надо для появления жизни?
«Первичный бульон» как среда для появления жизни имеет свои недостатки. В водной среде белки, РНК и ДНК неустойчивы. Эти длинные молекулы со временем распадаются на отдельные звенья – аминокислоты или нуклеотиды. Химическое соединение аминокислот в белок или нуклеотидов в РНК происходит с выделением воды. Поэтому когда ее вокруг много, равновесие этой реакции смещено в сторону распада белка или РНК (такой распад с участием воды называется «гидролиз»). Первым обратил внимание на эту проблему в контексте происхождения жизни Джон Бернал еще в 1949 году. Клетки строят длинные молекулы, используя активированные нуклеотиды и аминокислоты. При построении РНК и ДНК в ход идут нуклеотид-трифосфаты, их соединение в цепочку сопровождается выделением пирофосфорной кислоты, а не воды. Похожая хитрость позволяет клеткам собирать белки. Однако в аппарате Миллера получаются обычные, а не активированные аминокислоты. Без сложных клеточных систем активации получить из них белки в водной среде нельзя. Поэтому на безжизненной планете для соединения аминокислот и нуклеотидов в цепочки надо как-то избавляться от воды. Для этого хорошо подходят, например, заливы и лагуны по берегам океана, которые наполняются водой во время прилива и пересыхают в отлив. Похожие условия частого высыхания возможны в вулканических районах благодаря регулярным выбросам горячей воды из гейзеров. Эксперименты показали, что запекание сухих смесей нуклеотидов при температуре 120 °C приводит к образованию коротких цепочек РНК из 3–10 нуклеотидов. Для соединения аминокислот в белки эти условия слишком жесткие, они приводят к разложению самих аминокислот. Однако, как выяснилось, полного высыхания и не требуется: достаточно, чтобы в растворе не было «химически свободной» воды. В крепком рассоле (100 и более граммов NaCl на литр, как в Мертвом море) все молекулы воды прочно связаны с ионами натрия и хлора и не являются химически свободными. Поэтому в рассоле при температурах 60–80 °C равновесие смещено в сторону образования связей, и аминокислоты соединяются в короткие цепочки (эта реакция называется «солевой пептидный синтез» и будет подробнее описана в следующих главах). Так что, хотя вода и необходима для жизни, ее не должно быть слишком много. Другая проблема теории «первичного бульона» связана с тем, что в нем нет границ. В клетках тысячи генов «играют в одной команде» благодаря клеточной мембране, отделяющей их от внешней среды. Чтобы возникла простейшая кооперация между разными РНК, одни из которых занимаются копированием, а другие, например, готовят для этого детали-нуклеотиды, эти РНК должны как-то отделить свою тесную компанию от остального мира. Тогда нуклеотиды пойдут на копирование только тех РНК, которые их делали, а копирующая РНК будет копировать только себя и своих соседей, которые поставляют ей нуклеотиды. В безграничном бульоне же эти нуклеотиды будут расплываться и достанутся всем соседям, независимо от их вклада в общее дело. Копирующая РНК, которая попытается копировать не только себя, но и другие молекулы, в условиях бульона не сможет отличить союзников от прихлебателей и в итоге проиграет в конкуренции. Ученые предложили два разных решения проблемы границ, которые по аналогии с «первичным бульоном» были названы «первичная пицца» и «первичный майонез». В модели «первичной пиццы», изложенной в книге «The Major Transitions in Evolution» (John Maynard Smith, Eö rs Szathmá ry, 1995), предполагается, что жизнь зарождалась в виде тонкого слоя органических молекул на поверхности глины (например, каолинита или смектита), частицы которой состоят из тонких алюмосиликатных слоев. Глина может набухать, потому что молекулы воды, как и другие катионы и анионы, внедряются между этими слоями, увеличивая расстояния между ними. Эксперименты показали, что глина довольно прочно связывает аминокислоты, нуклеотиды, белки, РНК и другие биологические молекулы. На поверхности кристаллов глины и между ее слоями они накапливаются в высокой концентрации даже из очень разбавленного раствора. Адсорбирующие свойства глины используются и в быту: вы наверняка видели смектит под названием «минеральный наполнитель для кошачьих туалетов». На глине нуклеотиды самопроизвольно выстраиваются именно так, как нужно для их «сшивания» в цепочку РНК. Длинные молекулы РНК очень редко отделяются от глины полностью, но могут медленно перемещаться по ее поверхности, открепляясь то одним, то другим концом. Математическое моделирование взаимодействия разных РНК на плоской минеральной поверхности (Czaran T., Szathmary E., 2000) показало, что в таких условиях легко образуются группы разных молекул, связанных взаимной помощью, а размножение паразитов ограничено и не приводит к вымиранию кооперирующихся РНК. Так что, хотя четких границ на минеральной поверхности нет, она достаточно ограничивает подвижность РНК и нуклеотидов, чтобы могли появиться тесные группы взаимопомощи. Еще один вклад минеральной подложки в возникновение жизни состоит в том, что она может работать катализатором, т. е. ускорять химические реакции. Эта функция выходит на первый план в тех вариантах модели «первичной пиццы», в которых в качестве минеральной основы предлагается не глина, а сульфидные минералы – пирит (FeS2), сфалерит (ZnS), алабандин (MnS). Дальше в этой главе мы подробнее рассмотрим связь сульфидных минералов с биохимией. Модель «первичного майонеза» предложена Гарольдом Моровицем в книге «Mayonnaise and The Origin of Life: Thoughts of Minds and Molecules». Она предполагает, что примитивные аналоги клеточных мембран существовали с древнейших времен, еще до появления самокопирующихся РНК. Иными словами, весь мир РНК существовал внутри протоклеток – мелких жировых пузырьков. Теория «первичного майонеза» имеет меньше сторонников, чем теория «первичной пиццы», потому что для протоклеток существует проблема питания: нуклеотиды очень плохо проходят через мембраны. В современных клетках для этого существуют специальные транспортные белки, но адекватного решения для поглощения нуклеотидов примитивными протоклетками пока не найдено. Зато в модели «первичного майонеза» достигается очень эффективное разделение молекул РНК на кооперирующиеся группы, поэтому отвергать ее ученые не спешат. Более того, есть пути совмещения теорий «первичной пиццы» и «первичного майонеза»: частицы глины, как оказалось, помогают образованию мембранных пузырьков, при этом возникший пузырек окружает частицу глины со всех сторон. Солнце: друг или враг? Почти вся современная жизнь прямо или косвенно зависит от энергии солнечного света, которая в ходе фотосинтеза используется для построения сахаров и других клеточных веществ. Даже глубоководные сообщества, обитающие в полной темноте, зависят от кислорода, вырабатываемого водорослями в верхних слоях океана. Как известно, в стратосфере существует озоновый слой, который поглощает 99 % ультрафиолетового излучения Солнца. Озон (О3) образуется из кислорода под действием того же ультрафиолета. В древние геологические эпохи, когда кислорода в атмосфере еще не было, озонового слоя тоже не могло быть. Более того, молодое Солнце излучало больше ультрафиолета, чем сейчас, когда оно, разменяв пятый миллиард лет, стало спокойной звездой среднего возраста. Поэтому во времена появления жизни на Земле ее поверхность подвергалась мощному ультрафиолетовому излучению, и смертельная для многих современных организмов доза излучения набиралась за несколько минут. В связи с этим многие ученые склонялись к тому, чтобы спрятать первые живые организмы поглубже под воду для защиты от губительных лучей. Однако ультрафиолетовое излучение вызывает самые разнообразные химические реакции, в том числе ведущие к синтезу аминокислот и нуклеотидов из простых молекул. Так что для каких-то этапов происхождения жизни оно, наоборот, могло быть полезно. Как же разобраться, какие из первых шагов жизни происходили на свету, а какие – в темноте? Ответ на этот вопрос пришел с неожиданной стороны. В последние годы бурно развивается синтетическая биология, целью которой является создание организмов с принципиально новыми свойствами. Например, коллектив под руководством Стивена Беннера достиг больших успехов в создании альтернативных нуклеотидов. Эти искусственные звенья хорошо встраиваются в ДНК и РНК обычными природными ферментами, образуют комплементарные пары друг с другом, но не со стандартными нуклеотидами А, Г, Т и Ц, и расширяют нуклеотидный алфавит до шестибуквенного (рис. 6.1) (Malyshev et al., 2014; Yang et al., 2011 (русский краткий анонс: http: //www.chemport.ru/datenews.php? news=2557)).
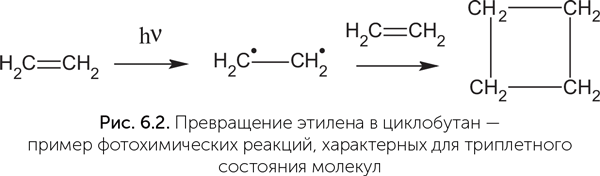
Получается, что с задачей хранения генетической информации в принципе могут справиться самые разные варианты нуклеиновых оснований, и А, Г, Т, Ц, возможно, были отобраны природой совсем по другим признакам. Как считает известный биофизик Армен Мулкиджанян, таким признаком была устойчивость к ультрафиолетовому излучению (Mulkidjanian, A. Y., Galperin, M. Y., 2007). Здесь надо пояснить, как устроены молекулы и как происходит их взаимодействие со светом. Каждая химическая связь, изображаемая в структурных формулах линией между атомами, обычно состоит из двух электронов, которые вместе движутся между двумя связанными атомами. У каждого электрона есть собственное магнитное поле, направление которого называется «спин». Два электрона, образующие химическую связь, имеют противоположные спины, так что их магнитные поля взаимно компенсируются. Такие электроны называются «спаренными». Если молекулу разорвать на две части, то электроны из разорванной связи имеют два варианта дальнейшей судьбы. Они могут разойтись по одному в каждый фрагмент молекулы или оба вместе в один из фрагментов. В первом случае эти электроны остаются без пары и готовы к образованию новой связи с любой подходящей молекулой. Фрагменты молекул, имеющие неспаренный электрон, очень химически активны и называются «радикалы». Во втором случае, когда одному фрагменту достаются два электрона, а другому – ни одного, эти фрагменты имеют электрический заряд и называются «ионами». Неспаренных электронов в них нет, и они более стабильны, чем радикалы. Когда в молекулу попадает фотон с подходящей энергией, он поглощается парой электронов, образующей химическую связь, и молекула переходит в возбужденное состояние с избыточной энергией. Возбужденных состояний как минимум два. Сначала молекула оказывается в неустойчивом и короткоживущем состоянии (так называемом синглетном состоянии). В нем спины электронов возбужденной пары еще антипараллельны, как и в спокойном состоянии молекулы. В синглетном состоянии молекула может сбросить возбуждение и вернуться в исходное состояние путем флюоресценции (излучения светового кванта с энергией чуть меньше исходной) или рассеяния энергии в тепло либо перейти в следующее – триплетное – состояние, в котором спины электронов становятся параллельными и химическая связь между атомами фактически разрывается. Если в молекуле была возбуждена одинарная связь, то молекула разрушается в этом месте. Если же была возбуждена двойная связь (точнее, так называемая пи-электронная система, образующая «вторые палочки» двойных связей), то молекула в триплетном состоянии сохраняет целостность, но становится бирадикалом – иными словами, у нее теперь имеются два неспаренных электрона, которые могут образовать две новые химические связи. Поэтому молекула в триплетном состоянии химически активна и вступает в разнообразные реакции. Например, молекулы этилена (С2H4), имеющие двойную связь между атомами углерода, при УФ-облучении частично объединяются попарно в циклобутан (С4H8), у которого вместо одной двойной связи образуются две одинарные связи между двумя дополнительными атомами углерода (рис. 6.2). Молекула может также вернуться из триплетного состояния в основное, невозбужденное путем излучения кванта света – фосфоресценции. В отличие от флюоресценции фосфоресценция может происходить спустя минуты и часы после облучения вещества, а разница в энергии поглощенного и излученного кванта света больше.
Так вот, у природных азотистых оснований синглетное состояние крайне короткоживущее. Оно легко рассеивает энергию возбуждения в тепло через колебания и вращение молекулы, обмен атомами водорода и другие механизмы и возвращается обратно в невозбужденное состояние. Синглетное состояние пуриновых оснований, аденина и гуанина, живет около 10–12 секунды – примерно в 10 000 раз меньше, чем синглетные состояния большинства молекул сравнимого размера и сложности, например аминокислоты триптофана. Благодаря быстрому рассеиванию энергии они из синглетного состояния практически всегда переходят в невозбужденное, а не в химически активное триплетное. А раз азотистые основания практически не попадают в триплетное состояние, то и разрушение их под действием ультрафиолета происходит очень редко. Пиримидиновые основания, цитозин и тимин, рассеивают энергию несколько хуже, чем пурины, и, соответственно, менее устойчивы. Однако образование комплементарных пар улучшает рассеивание энергии еще примерно в 50 раз благодаря обмену протонами в водородных связях пары. Поэтому устойчивость комплементарной пары нуклеотидов к ультрафиолету выше, чем каждого из них по отдельности. Кроме того, в нуклеиновых кислотах плоские молекулы азотистых оснований лежат стопкой, поэтому их пи-электронные системы взаимодействуют между собой (так называемое стэкинг-взаимодействие) и могут передавать друг другу энергию возбуждения, еще усиливая рассеивание и дополнительно увеличивая устойчивость к ультрафиолету – до 20 раз по сравнению с одной комплементарной парой нуклеотидов (Mulkidjanian et al., 2003). Азотистые основания не только сами устойчивы к ультрафиолету, они защищают соседние молекулы. Например, они предохраняют от УФ-расщепления фосфоэфирную связь (О-Р). При облучении УФ глицеролфосфата отщепление фосфорной кислоты происходит в 300 раз быстрее, чем при облучении аденозинмонофосфата, а образование комплементарных пар и стэкинг-взаимодействие в цепочке ДНК или РНК еще увеличивают степень защиты. Таким образом, солнечный ультрафиолет, который в принципе разрушительно воздействует на нуклеотидную цепочку, как и на любые органические молекулы, может служить фактором отбора по следующим направлениям: • отбор самых УФ-стойких азотистых оснований; • отбор азотистых оснований, склонных образовывать комплементарные пары; • отбор нуклеотидов одной хиральности из смеси правых и левых нуклеотидов (так называемой рацемической смеси), потому что смесь правых и левых нуклеотидов в цепочке нарушает стэкинг-взаимодействие; • отбор длинных молекул РНК по сравнению с более короткими, потому что в длинных цепочках стэкинг-взаимодействие увеличивает устойчивость к ультрафиолету; • отбор молекул РНК, содержащих двуспиральные участки (шпильки), среди молекул со случайными последовательностями, потому что в них больше нуклеотидов входят в состав комплементарных пар. |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 65; Нарушение авторского права страницы