|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
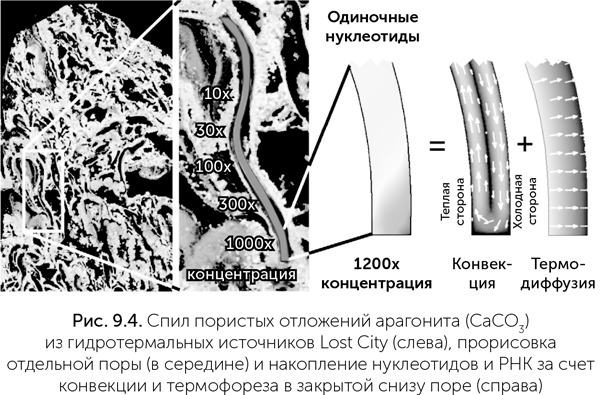
Копирование РНК в тепловой ловушке
Все процессы соединения нуклеотидов в РНК очень чувствительны к концентрации нуклеотидов, которые в разбавленном растворе гораздо хуже соединяются в цепочки. К сожалению, все известные пути получения нуклеотидов, возможные в природных условиях, дают относительно разбавленные растворы продуктов. Было бы очень хорошо найти какой-нибудь эффективный механизм их концентрирования. Как мы помним, возникновение жизни произошло на горячих источниках. В этом ландшафте возникают пересыхающие лужи, в которых могут накапливаться различные растворенные вещества, например формамид. К сожалению, для накопления нуклеотидов этот механизм не очень подходит: при испарении воды увеличивается концентрация всех растворенных веществ, включая минеральные соли, которых заведомо больше, чем нуклеотидов. Но, как оказалось, на тех же геотермальных полях есть механизмы концентрирования, которые отличают крупные молекулы нуклеотидов и РНК от растворенных минеральных солей. Для геотермальных полей характерны мелкопористые осадки и устойчивая разница температуры между горячей подземной водой и холодным воздухом. В этих условиях в порах и трещинах, заполненных водой, возникает устойчивая конвекция: поток жидкости и тепла. Благодаря конвекции и перепадам температур эти трещины становятся удобным местом концентрирования нуклеотидов и копирования РНК. В работе Baaske et al. (2007) изучалось поведение нуклеотидов и РНК разной длины в поре, закрытой снизу и открытой сверху в холодную воду. При подогреве сбоку в такой поре происходят конвекция жидкости и перенос растворенных молекул вдоль градиента температуры (термофорез). Оказалось, что нуклеотиды и РНК в такой поре подсасываются из холодной воды и эффективно накапливаются в нижней части холодной стенки (рис. 9.4). Для поры шириной 1 мм и длиной 10 мм получается концентрирование нуклеотидов и коротких РНК в шесть-семь раз. Но степень концентрирования очень сильно (экспоненциально) зависит от отношения длины к ширине поры. Пора размером 0, 1 мм × 10 мм или 1 мм × 100 мм концентрирует нуклеотиды примерно в 100 млн раз. РНК длиной примерно 40 нуклеотидов и более ведут себя по-другому по сравнению с одиночными нуклеотидами и короткими РНК. Они очень сильно концентрируются даже в коротких порах – в 20 000 раз в поре 1 мм × 10 мм для РНК длиной 100 нуклеотидов. Длинные молекулы РНК, попавшие в такую пору, практически неизбежно захватываются ею и накапливаются на маленьком (меньше 0, 01 кв. мм) участке в нижней части холодной стенки. Концентрация РНК там ограничена только их физическими размерами.
Если в такой закрытой нагреваемой поре происходит еще и образование РНК из нуклеотидов (здесь неважно, идет ли речь о копировании существующей РНК или о случайной сборке), то конвекция, термофорез и накопление нуклеотидов очень способствуют образованию длинных РНК. В работе Маста с коллегами (Mast et al., 2013) было рассчитано, что в тепловой ловушке будут получаться РНК длиной 200–300 нуклеотидов безо всяких ферментов и рибозимов. К сожалению, по техническим причинам проверяли они эти расчеты не на полимеризации РНК из отдельных нуклеотидов, а на ДНК, и при этом еще не сборка отдельных нуклеотидов, а стыковка 95-нуклеотидных фрагментов ДНК при помощи одноцепочечных «липких концов». Такие видоизменения потребовались потому, что только к фрагментам ДНК такой длины возможно присоединить флуоресцентные метки, не мешающие полимеризации. Но зато благодаря таким меткам удалось в реальном времени и с высоким разрешением измерить накопление и полимеризацию ДНК на нижнем конце холодной стенки. И в точном соответствии с теорией получилось, что молекулы ДНК в этих условиях получаются в 10–20 раз длиннее, чем при любой одинаковой со всех сторон температуре. В поре, открытой с обоих концов, поведение молекул несколько отличается. Если есть проток жидкости снизу вверх и нагрев с одной стороны, то в поре тоже происходят конвекция и термофорез. Молекулы РНК накапливаются на нижнем конце холодной стенки, но их поведение сильно зависит от длины. Нуклеотиды и короткие РНК слабее увлекаются термофорезом в нисходящий холодный поток и в итоге вымываются из поры с восходящим потоком, а РНК длиннее определенного порога (он зависит от скорости потока и размеров поры) накапливаются в ней. Это избирательное удержание длинных РНК может противостоять быстрому размножению коротких РНК и сохранять длинные молекулы в ряду поколений несмотря на то, что они копируются медленнее (Kreysing et al., 2015). И в открытых, и в закрытых порах благодаря конвекции молекулы РНК последовательно попадают в горячие и холодные зоны. В горячей зоне двухцепочечная РНК может разделяться на одиночные цепи, которые смогут стать матрицами для следующего цикла копирования. Мир шпилек Изящное решение проблемы затравок было недавно предложено Александром Марковым (http: //macroevolution.livejournal.com/170098.html). Оно состоит в том, что в условиях неферментативного копирования РНК преимущества получают те последовательности, которые могут служить затравками для синтеза собственных копий (рис. 9.5). Такими свойствами обладают палиндромные последовательности РНК. Палиндромами называли слова или фразы, которые читаются одинаково в обе стороны («А роза упала на лапу Азора»). В случае РНК и ДНК палиндромной последовательностью называют такую, которая читается одинаково слева направо и – после замены нуклеотидов на комплементарные – справа налево. Например, последовательность GGACCUAGGUCC будет палиндромом.
При случайном разрыве таких цепей РНК получаются короткие фрагменты, которые тоже могут служить затравками для удлинения своих родственников. В условиях неферментативного копирования РНК идея Маркова («Мир палиндромов») превращает необходимость затравок из проблемы в фактор естественного отбора, который начинается еще до появления первого активного рибозима, с молекулами РНК длиной 15–20 нуклеотидов. Причем низкая скорость неферментативного копирования здесь не страшна: достройка концов палиндромных РНК требует присоединения лишь 5–10 нуклеотидов за один цикл. Палиндромность первых РНК повышает вероятность появления рибозимов, так как палиндромные последовательности образуют множество шпилек (рис. 9.6).
Следы такого способа роста структур РНК до сих пор видны в древнейшем реликте РНК-мира – молекулах транспортных РНК (тРНК). Эти молекулы длиной 76 нуклеотидов имеют форму клеверного листочка. «Стебелек» и каждый из «листиков» трилистника включают двуспиральные участки РНК. Три «листика» образуют шпильки, а стебелек заканчивается свободными концами цепи РНК. В последовательности тРНК давно были обнаружены внутренние повторы, допускающие другие варианты укладки молекулы, например, в виде одной длинной двухцепочечной шпильки с несколькими внутренними петлями. На основе этих повторов были предложены модели роста тРНК путем увеличения количества палиндромных сегментов молекулы (Rodin et al., 2011; DiGiulio, 2009). Если у нас есть механизм абиогенного копирования РНК, хотя бы только коротких и палиндромных молекул, то среди них могут оказаться РНК с какой-то рибозимной активностью. Строго говоря, не только лигаза и полимераза способны ускорить накопление своих копий в такой системе. Например, рибозимы-экзонуклеазы, разрушающие молекулы РНК, могут быть полезными для их точного и быстрого копирования. Экзонуклеазы работают в клетках вместе с полимеразами и служат для отрезания неправильно присоединенных нуклеотидов, повышая точность копирования. Рибозим с такой активностью мог бы повышать точность неферментативного копирования РНК и косвенно – скорость копирования, потому что после неправильно присоединенного нуклеотида (не образующего комплементарную пару с матрицей) следующий нуклеотид присоединяется намного медленнее обычного. Кроме того, для увеличения количества собственных копий рибозим может вообще воздействовать не на копирование РНК, а на предшествующие шаги – превращение простых органических веществ в нуклеотиды. Иначе говоря, появляется обмен веществ – химические реакции малых молекул, происходящие под контролем ферментов. В следующей главе мы подробнее рассмотрим, на что способны рибозимы в процессе обмена веществ.
Глава 10 |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 60; Нарушение авторского права страницы