|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Модульная структура окислительно-восстановительных ферментов
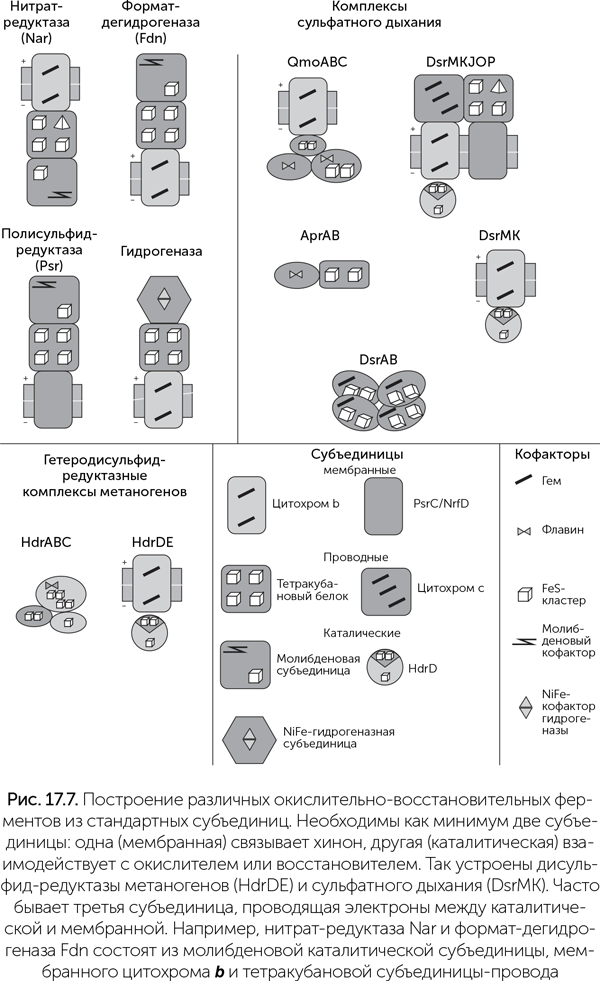
Изучение большого разнообразия дыхательных цепей показывает, что множество ферментных комплексов, работающих с разными веществами, строятся из небольшого набора субъединиц, как в конструкторе LEGO (Baymann et al., 2003). В простейшем случае дыхательная цепь состоит из двух комплексов, перенос электронов между которыми осуществляется хиноном. Каждый из двух комплексов включает как минимум три блока: • каталитическая субъединица получает электроны от восстановителя; • мембранная субъединица отдает электроны на хинон; • соединительная субъединица работает «проводом» между двумя другими. Среди каталитических субъединиц выделяются своей универсальностью молибденовые оксидоредуктазы (их еще называют CISM – Complex Iron-Sulfur Molibden). Это семейство ферментов восстанавливает гидрокарбонат (до муравьиной кислоты), нитрат (до нитрита), хлорат, перхлорат, арсенат, селенат, полисульфиды, диметилсульфоксид, триметиламиноксид и даже хлорорганические вещества. В состав этой субъединицы входит молибден, связанный с двумя молекулами витамина В9 (этот комплекс называется «молибденовый кофактор», MoCo), и один 4Fe-4S кластер (рис. 17.7). Гетеродисульфид-редуктазная субъединица содержит три железосерных кластера 4Fe-4S (один из них связан с белком через пять атомов серы, а не через четыре, как обычно). Она входит в состав нескольких ферментов метаногенеза и сульфатного дыхания, у метаногенов восстанавливает дисульфидный мостик между двумя коферментами (CoB-S-S-CoM), у сульфатредукторов – мостик между двумя цистеинами в переносящем серу белке DsrC. NiFe-гидрогеназная субъединица работает с молекулярным водородом (Н2). Она входит в состав разных гидрогеназ, мембранных и растворимых, поглощающих и выделяющих водород, а также формат-гидрогенлиазы и даже комплекса I (НАДН-хинон-оксидоредуктазы), и сочетается с разными «проводами» и хинонными портами.
Трансмембранные цитохромы b сочетают функции «проводов» и «хинонных разъемов» и используются, когда каталитическая субъединица должна быть по одну сторону мембраны, а хинон – по другую. Они содержат два гема b и входят в состав формат-дегидрогеназы, нитрат-редуктазы, гидрогеназ, комплекса b 6 f в фотосинтезе и многих других ферментов. Другой белковый модуль с хинонным сайтом носит обозначение PsrC/NrfD. Эта субъединица связывает каталитический центр с хинонами на той же стороне мембраны, в отличие от цитохромов b. Она входит в состав полисульфид-редуктаз Psr, арсенат-редуктазы, аммонифицицирующей нитрит-редуктазы Nrf и некоторых гидрогеназ. В составе некоторых комплексов эта субъединица дополнительно переносит протоны через мембрану за счет изменения формы белка. Кроме трансмембранных «проводов» в комплексах электрон-транспортной цепи часто бывают гидрофильные провода, соединяющие разные субъединицы. Основные типы гидрофильных проводов – это тетракубановый белок (4 кластера 4Fe-4S), бактериальный ферредоксин (2 кластера 4Fe-4S, более известен как подвижный переносчик) и двухгемовый цитохром с. Например, формат-дегидрогеназа Fdn состоит из трех субъединиц: молибденовой каталитической, тетракубанового «провода» и хинон-связывающего цитохрома b. Нитрат-редуктаза Nar собрана из таких же модулей, но в другой ориентации – ее каталитический домен обращен внутрь клетки, а не наружу. Ферменты сульфатного дыхания дают примеры более сложных структур. Например, комплекс Qmo, проводящий электроны к аденозил-фосфосульфат-редуктазе (Apr), занимается сведением в один ток электронов от хинона и ферредоксина. В его составе есть цитохром b, дикубановая субъединица и две флавин-содержащие субъединицы. Комплекс DsrMKJOP, восстанавливающий переносчик серы DsrC, состоит обычно из пяти субъединиц: каталитической гетеродисульфид-редуктазной, двух мембранных хинонных портов (цитохром b и PsrC/NrfD типа) и двух вспомогательных на наружной стороне мембраны (трехгемовый цитохром с и тетракубановая). Зачем такая сложность – непонятно, потому что в некоторых клетках с той же работой справляется комплекс DsrMK из двух субъединиц – одной каталитической и одного цитохрома b. В группе дельта-протеобактерий ферменты сульфатного дыхания и другие комплексы еще более усложнены и содержат по несколько субъединиц цитохромов с, которые могут иметь до четырех-шести гемов. Видимо, это позволяет им быстро переключаться между разными окислителями и восстановителями. Происхождение комплекса I (НАДН-хинон-оксидоредуктаза) Один из сложнейших комплексов дыхательной цепи митохондрий и многих бактерий, комплекс I, переносит электроны с НАДН на хиноны. Энергия этих электронов используется для откачки из клетки протонов, которые затем входят обратно через роторную АТФазу и производят АТФ. У архей Methanosarcinales есть родственные комплексы, переносящие электроны с водорастворимого кофермента F420 на мембранный аналог хинона, метанофеназин. Минимальный вариант комплекса I включает в себя 14 разных субъединиц. В митохондриях животных комплекс I еще более сложен, и количество его субъединиц может превышать 40. Комплекс I по форме похож на букву Г, одно плечо которой лежит в мембране, а другое выступает в цитоплазму. Сайт связывания НАДН находится на конце цитоплазматического плеча. Четыре из мембранных субъединиц (J, L, M, N) непосредственно переносят протоны через мембрану за счет изменения формы белка (рис. 17.8).
Сравнение аминокислотных последовательностей субъединиц комплекса I показало, что у многих из них есть родственники в других типах белков, прежде всего среди гидрогеназ и натрий-протонных антипортеров[19]. Гидрогеназы – это разнообразная группа ферментов, работающих с молекулярным водородом. Они могут как вырабатывать водород, окисляя НАДН, ферредоксин и другие переносчики электронов, так и поглощать водород с восстановлением переносчиков. Эти функции востребованы, например, у организмов, использующих молекулярный водород для получения энергии и восстановления СО2 (ацетогены, метаногены и многие другие). По используемым металлам в активном центре гидрогеназы делятся на NiFe и FeFe семейства, которые дальше подразделяются на группы по составу субъединиц. Родством с комплексом I обладает четвертая группа NiFe гидрогеназ. В четвертой группе гидрогеназ наиболее просто устроены гидрогеназы Ech. Это мембранные ферменты, получающие электроны от цитоплазматических доноров и выделяющие водород. Они состоят из шести субъединиц, все они имеют гомологи в комплексе I. Четыре цитоплазматических субъединицы гидрогеназы Ech родственны субъединицам B, C, D, I комплекса I, а две мембранных – субъединицам H и L соответственно (рис. 17.8). Каталитический центр гидрогеназы Ech, содержащий по одному атому железа и никеля, расположен на гомологе D-субъединицы комплекса I. Более сложные гидрогеназы этой группы имеют дополнительные субъединицы. Например, Hyc – формат-гидроген-лиаза кишечной палочки – осуществляет разложение муравьиной кислоты с выделением СО2 и водорода. Она состоит из восьми субъединиц, шесть из которых похожи на субъединицы Ech, а две другие – молибден-содержащая формат-дегидрогеназа и субъединица, участвующая в передаче электронов от молибденового центра к никель-железному. Эти две субъединицы гомологичны, соответственно, С– и N-концам субъединицы G комплекса I. Гидрогеназа Hyf, тоже обнаруженная у кишечной палочки, имеет также гомологи мембранных субъединиц K, M, N комплекса I, которые ответственны за перенос протонов через мембрану. К сожалению, источник электронов для Hyf пока не найден. У других бактерий и архей обнаружены гидрогеназы четвертой группы, содержащие до 13 субъединиц и способные переносить ионы через мембраны. Для некоторых из них известны доноры электронов. Например, гидрогеназа Coo бактерий Rhodospirillum и Carboxydothermus окисляет угарный газ с выделением водорода и запасанием энергии. Известны также близкородственные гидрогеназам белки, не имеющие собственно гидрогеназного центра. Наиболее просто устроены комплексы Ehr, состоящие из двух цитоплазматических субъединиц (гомологи B и D комплекса I) и четырех находящихся в мембране, которые соответствуют субъединицам H, L, M и С-концу K в составе комплекса I. Их функции неизвестны. Более сложный гидрогеназоподобный комплекс Mbx архей Pyrococcus и Thermococcus содержит также гомологи субъединиц C и I и дополнительные мембранные субъединицы. Он переносит электроны с ферредоксина на НАДФ и сопрягает эту реакцию с переносом протонов через мембрану. Субъединицы L, M и N комплекса I ответственны за транспорт ионов. Помимо гидрогеназ четвертой группы родственные им белки входят в состав натрий-протонных антипортеров. Эти мембранные транспортные белки обменивают один ион натрия на один протон, перенося их через мембрану навстречу друг другу, и могут работать в обоих направлениях. Перенос ионов происходит благодаря движениям всей белковой молекулы. Эти антипортеры используются для соединения натриевой и протонной энергетики в одной клетке и могли быть одной из первых систем откачки протонов из клетки на заре протонной энергетики. Детальное изучение трехмерной структуры комплекса I показало, что субъединица H обладает сходной пространственной укладкой с L, M и N, но у нее отсутствуют аминокислоты, необходимые для связывания ионов. Видимо, она, подобно L, M и N, тоже происходит от натрий-протонных антипортеров, но в составе комплекса I изменила функцию и участвует в передаче движения от цитоплазматического плеча «рычага» к ионным насосам L – M-N. Мы видим, что шире всего распространены четыре субъединицы: мембранные H и L и цитоплазматические B и D. Они присутствуют в комплексе I, во всех мембранных гидрогеназах и гидрогеназоподобных комплексах, например Ehr. Поскольку они всегда оказываются в середине ферментных комплексов, их еще называют «универсальным адаптером». Субъединицы B и D также родственны двум субъединицам растворимых NiFe-гидрогеназ. Видимо, движение электронов в этих гидрогеназах сопровождалось изменением формы белка, и соединение B-D субъединиц с мембранными H-L, происходящими от натрий-протонного антипортера, позволило гидрогеназе запасать энергию, перенося ионы через мембрану. На следующем шаге по пути к комплексу I гидрогеназа, видимо, стала формат-гидрогенлиазой, подобно Hyc кишечной палочки. Для этого к комплексу присоединились две субъединицы, которые в современном комплексе I слились в единую субъединицу G. Потом к комплексу присоединились субъединицы E и F, родственные растворимым НАДН-дегидрогеназам, и донором электронов стала НАДН вместо муравьиной кислоты. Однако субъединица G, утратившая аминокислоты, нужные для связывания молибдена, до сих пор сохраняет сходство с формат-дегидрогеназами. Приблизительно тогда же субъединица D утратила сайт связывания никель-железного кластера и способность выделять водород, и акцептором электронов стал хинон. Разница редокс-потенциала между НАДН и хиноном больше, чем между муравьиной кислотой и водородом. Поэтому перенос электронов с НАДН на хинон позволяет получить больше энергии. Чтобы полнее использовать эту энергию, в результате дупликации субъединицы L возникли дополнительные ионные насосы M и N. Малая субъединица K похожа на вспомогательные субъединицы натрий-протонных антипортеров и, видимо, была позаимствована из них для помощи в работе L – M-N. Эволюция комплекса I дает нам яркий пример смены функций на молекулярном уровне. Миллиарды лет назад субъединицы комплекса I потеряли железно-никелевый гидрогеназный центр и молибденовый формат-дегидрогеназный центр. Однако субъединицы D и G не были утрачены из состава комплекса и до сих пор сохраняют сходство с соответствующими активными ферментами. Видимо, они встроены в передачу механического движения внутри комплекса, и их утрата приведет к неработоспособности всей большой молекулярной машины. Происхождение цитохромных комплексов bc 1 (комплекс III) и b 6 f Цитохромные комплексы bc 1 и b 6 f входят в состав электрон-транспортных цепей митохондрий, хлоропластов, цианобактерий и многих аэробных бактерий. Как можно догадаться из их названия, они состоят из нескольких цитохромов разных типов. Цитохромные комплексы переносят электроны с хинонов на водорастворимые высокопотенциальные переносчики, обычно малые цитохромы с или пластоцианины. Важной функцией этих комплексов является бифуркация, или разветвление пути электронов (процесс, обратный конфуркации электронов в комплексе Qmo сульфатного дыхания). Приняв два электрона с хинона на наружной стороне мембраны и выбросив наружу два протона, цитохромный комплекс пускает их по разным путям. Один электрон проходит к сайту растворимого переносчика (например, цитохрома с ) и следует дальше по электрон-транспортной цепи, а другой электрон попадает на внутреннюю сторону мембраны, где находится второй хинонный сайт комплекса bc 1. Когда хинон во втором сайте примет два электрона, он проходит через мембрану к первому хинонному сайту комплекса и отдает там электроны на цитохром и протоны в воду. За счет такого сложного пути электронов удается полнее использовать разницу потенциалов между хиноном и цитохромом с: на каждый прошедший через комплекс электрон через мембрану переносятся два протона, а не один, как в простейшем Q-цикле.
Цитохромные комплексы bc 1 содержат от 3 субъединиц у ряда бактерий до 11 субъединиц в митохондриях. Три основные субъединицы, которые есть в любых вариантах комплекса bc 1, – это трансмембранный цитохром b (8 трансмембранных спиралей, два гема b ), цитохром с и железосерный белок Риске. Последний содержит необычный железосерный кластер 2Fe-2S, связанный с двумя цистеинами и двумя гистидинами. Из-за такого аминокислотного окружения его редокс-потенциал гораздо выше, чем у ферредоксинов, в которых железосерные кластеры связаны с белком через четыре цистеина. Комплексы b 6 f устроены сходно с bc 1, но вместо одного цитохрома b в них есть два разных белка: цитохром b 6 с четырьмя трансмембранными спиралями и белок PetD с тремя спиралями. Они по последовательности аминокислот сходны с N– и C-концевыми половинами цитохрома b комплекса bc 1, но связывают больше кофакторов: два гема b на цитохроме b 6 и по одной молекуле гема с, хлорофилла а и каротина на белке PetD. Цитохром f функционально аналогичен цитохрому c 1 комплекса bc 1 и тоже содержит гем c, но совсем не похож на цитохром c 1 по последовательности и имеет другое происхождение. Белки Риске комплексов bc 1 и b 6 f устроены практически одинаково. История цитохромных комплексов запутана из-за горизонтальных переносов генов. Комплексы bc 1 -типа (с длинным цитохромом b ) встречаются как у бактерий, так и у архей, а комплексы типа b 6 f – только у бактерий. У некоторых бактерий в геноме закодировано до четырех разных комплексов, причем на филогенетическом дереве они находятся на различных ветвях. В одном геноме могут сочетаться гены комплексов bc 1 и b 6 f. Лишь недавно, с накоплением данных по большому количеству геномов, удалось установить, что на филогенетическом дереве есть несколько ветвей bc 1 комплексов и несколько ветвей b 6 f комплексов (Dibrova et al., 2013). Иными словами, переход от одного типа к другому – будь то слияние двух генов в один длинный цитохром b или разделение гена на два – происходил несколько раз. Это позволяет предположить вероятное направление эволюции: несколько независимых событий разделения гена в одной и той же точке, да еще с последующим появлением сайта связывания гема c в одном месте в разных ветвях комплексов b 6 f очень маловероятны. А эволюция в обратном направлении – слияние двух соседних генов одного оперона и утрата сайта связывания гема с – вполне могла происходить много раз независимо. Длинная 8-спиральная форма цитохрома b уникальна для комплекса bc 1. Короткие 4-спиральные цитохромы b встречаются, помимо комплекса b 6 f, еще в ряде мембранных окислительно-восстановительных ферментов, например в формат-дегидрогеназе, и участвуют в проведении электронов через мембрану. Такие белки могли быть востребованы еще во времена LUCA, чтобы снимать электроны с внеклеточных доноров (например, кристаллов сульфида цинка) и использовать их для восстановления веществ внутри клетки. Функция современных комплексов bc 1 и b 6 f – разветвление электронов – могла возникнуть только с появлением достаточно сильных окислителей. Появление кислородного фотосинтеза сделало такой сильный окислитель (кислород) доступным для биосферы, но до того единственным сильным окислителем были хлорофиллсодержащие фотосистемы. Большинство комплексов b 6 f типа встречаются у фотосинтетиков (цианобактерий, Chlorobi и гелиобактерий) и участвуют в фотосинтезе. Хлорофилл и каротин в составе комплексов b 6 f участвуют в регуляции транспорта электронов между фотосистемами в зависимости от освещенности. Так что, скорее всего, первые комплексы b 6 f появились у процианобактерий путем объединения цитохрома b, белка Риске (он присутствует и в других редокс-ферментах, в том числе древних) и других субъединиц. Дальше они распространялись путем горизонтального переноса, в том числе вместе с генами фотосинтеза: у гелиобактерий все субъединицы b 6 f комплекса входят в супероперон фотосинтетических генов. У нефотосинтезирующих клеток светозависимая регуляция комплекса b 6 f не требовалась, хлорофилл и каротин терялись, и слияние субъединицы PetD с 4-спиральным цитохромом b 6 помогало стабилизировать структуру комплексов после потери этих кофакторов. Дальнейшая эволюция цитохромных комплексов шла под давлением кислорода: они были оптимизированы для уменьшения образования повреждающих активных форм кислорода и приспособлены к работе в дыхательной цепи аэробного дыхания. |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 118; Нарушение авторского права страницы