|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Происхождение цитохром-с-оксидазы
Ключевой фермент аэробного дыхания, комплекс IV, или цитохром-с-оксидаза, завершает дыхательную цепь и переносит электроны с цитохрома с на кислород. Она относится к семейству гем-медных оксидаз (НСО, haem-copper oxydase). Разные ферменты этого семейства восстанавливают кислород до воды либо оксид азота NO до закиси азота N2O. Все основные функции этих ферментов выполняются одной большой субъединицей, содержащей 12 трансмембранных спиралей, два гема и атом меди между тремя гистидинами (рис. 17.10). У работающих с NO ферментов этого семейства вместо атома меди присутствует железо. По сходству последовательностей главной субъединицы и набору вспомогательных субъединиц семейство делят на четыре подсемейства: НСО-А, НСО-В и НСО-С, которые восстанавливают кислород и переносят протоны через мембрану, и NOR, который восстанавливает NO и не переносит протоны. Подсемейство НСО-А, к которому относится и комплекс IV митохондрий (см. главу 15), оптимизировано для высоких концентраций кислорода и переносит больше протонов, чем НСО-В и НСО-С. Среди ученых существуют очень разные точки зрения о происхождении и эволюции этого семейства ферментов. Например, в работах группы Анны-Лизы Дуклузье и Вольфганга Ницшке в Марселе (Ducluzeau et al., 2009) отстаивается сценарий, в котором ферменты подсемейства NOR (переносящие электроны с хинона на NO) были еще у LUCA и участвовали в нитритном дыхании или защите от оксида азота, а кислородные подсемейства произошли от него после появления кислородного фотосинтеза минимум два раза независимо. В пользу этого сценария ученые приводят филогенетические деревья, на которых подсемейство NOR, в отличие от других, четко делится на архейную и бактериальные ветви с корнем между ними. Именно такое дерево мы можем ожидать для белка, который был унаследован первыми бактериями и археями прямо от LUCA, а не распространялся путем горизонтального переноса. Кроме того, до появления кислорода на Земле вся медь была связана в нерастворимых минералах в одновалентном состоянии, например в Cu2S, и не использовалась клетками. Только с появлением кислорода эти минералы стали окисляться в относительно растворимые соединения двухвалентной меди, такие как CuSO4. Все медьсодержащие ферменты эволюционно относительно молоды, и железо в активном центре NOR, казалось бы, свидетельствует о древности этого подсемейства по сравнению с медь-содержащими HCO-A, B и C. Остаток тирозина, абсолютно необходимый для восстановления кислорода, находится в HCO-C совсем не там, где в HCO-A и НСО-В. Следовательно, переход этих ферментов на работу с кислородом был вызван разными, независимыми мутациями, добавившими тирозин к активному центру. Другие ученые, например Грибальдо (Gribaldo et al., 2009), используя несколько другие методы построения деревьев, обращают внимание на то, что НСО-А распространены в очень многих группах бактерий и архей, тогда как другие подсемейства ограничены в распространении. На основании их деревьев получается, что подсемейство НСО-В появилось у архей Sulfolobales и попало к бактериям путем горизонтального переноса, а НСО-С и NOR – изобретение протеобактерий. Дерево подсемейства НСО-А в их работе очень похоже на дерево 16S рибосомных РНК, не подверженных горизонтальному переносу, и предполагается, что НСО-А еще во времена LUCA участвовало в защите от кислорода – правда, непонятно, откуда кислород мог тогда взяться. Некоторые факты не укладываются ни в один из этих двух сценариев: например, в составе вспомогательных субъединиц NOR есть медь, значит, и это подсемейство должно быть не старше, чем кислородный фотосинтез. Другой факт связан с липидами. Известно, что для работы НСО во впадинах главной субъединицы должны быть связаны молекулы липидов бактериального типа – с жирными кислотами. Чтобы HCO работал в мембране археи, состоящей из терпеноидных липидов (см. главу 15), в ней должна быть небольшая примесь липидов бактериального типа. Археи, использующие НСО, всегда имеют также ферменты для синтеза бактериальных липидов, явно полученные горизонтальным переносом от бактерий (Dibrova et al., 2014). Иными словами, все НСО архей должны быть получены от бактерий, а не унаследованы от LUCA. При таком переносе мембранный белок оказывается в новом липидном окружении, к которому он не был приспособлен. Оптимизация перенесенного белка для работы в окружении архейных липидов приводит к ускоренной эволюции его последовательности и ошибкам при построении филогенетических деревьев. Прорыв в понимании происхождения этого семейства наметился в 2014 году. Биоинформатики из Техасского университета, используя чувствительные методы поиска сходства белков, нашли дальних родственников семейства НСО (Pei et al., 2014). Молекула НСО обладает несовершенной трехлучевой симметрией: 12 трансмембранных спиралей образуют 3 похожие группы по 4 спирали, расположенные вокруг общего центра. Два гема и один атом меди нарушают симметрию. Среди обнаруженных родственников НСО часть белков имеет 4 трансмембранные спирали, похожие больше всего на 9–12-ю спирали НСО (их назвали НСОН-s, НСО homolog single domain). Такие белки не могут устойчиво свернуться поодиночке, а должны объединяться по три, чтобы получилась трехмерная укладка, похожая на HCO. Если в геноме есть несколько генов HCOH-s, кодируемые ими белки могут объединяться в комплексы смешанного состава из двух субъединиц одного типа и одной – другого. Судя по наличию связывающих гем остатков гистидина, комплексы из одинаковых субъединиц могут содержать три молекулы гема, а комплексы состава «2+1» – одну. Другие родственники НСО, НСОН-t (НСО homolog triple domain) состоят из 12 спиралей, уложенных так же, как в НСО. Большинство из них связывают один гем, некоторые – два (рис. 17.10).
Почти все эти белки известны только из последовательностей полных геномов различных бактерий и никогда не изучались экспериментаторами. Только одна из семи групп HCOH-t попадала в руки экспериментаторов раньше. Это белок NnrS, выделенный из холерных вибрионов. У холерного вибриона он обеспечивает устойчивость к оксиду азота, вырабатываемому иммунной системой хозяина. У почвенных бактерий, где белок NnrS тоже был опознан, он организует движение клеток в сторону большей концентрации нитратов и нитритов. Точный механизм его работы неизвестен, в пробирке NnrS холерного вибриона не окисляет и не восстанавливает NO. Гены nnrS и большинства других родственников НСО в геномах соседствуют с генами ферментов нитратного и нитритного дыхания и генами защиты от отравления NO (не только иммунная система животных, но и сами бактерии травят им друг друга). Так что их функции должны быть как-то связаны с оксидами азота. 4-спиральные НСОН-s явно имеют отношение к предкам НСО, у которых еще не произошло слияния трех белковых субъединиц в одну. Остается ждать, пока биологи-экспериментаторы исследуют разведанные биоинформатиками цели, и тогда мы сможем судить о функциях предков гем-медных оксидаз.
Эволюция фотосистем Происхождение фотосистем от простых хлорофилл-связывающих белков с функцией защиты от ультрафиолета не вызывает больших сомнений. Гораздо менее понятно, как появилось два типа фотосистем. Существует две точки зрения. По одной гипотезе (слияния), ФСI и ФСII независимо возникли из светозащитных белков в разных линиях бактерий. В этом случае цианобактерии, имеющие оба типа фотосистем в одной клетке, появились благодаря событию горизонтального переноса генов одной из фотосистем. Другая гипотеза предполагает, что две фотосистемы возникли путем дупликации генов в одной клетке и появления какого-то «разделения труда» между копиями предкового гена. От этой клетки произошли цианобактерии, а потом вторая фотосистема распространилась к другим группам бактерий путем горизонтального переноса генов. Некоторые указания на порядок появления разных систем фотосинтеза можно найти в устройстве фотосистем. Так, реакционные центры фотосистем обычно состоят из двух белковых субъединиц. Это разные, хотя и родственные белки, возникшие в результате дупликации общего предкового гена. Однако у Chlorobi и гелиобактерий РЦ1 состоит из двух одинаковых субъединиц, т. е. их фотосистемы сохранили предковое состояние, существовавшее до дупликации. РЦ2 пурпурных бактерий и Chloroflexi состоят из двух разных субъединиц, как и ФСII цианобактерий. На филогенетическом дереве видно, что разные субъединицы РЦ2 пурпурных бактерий ближе друг к другу, чем к субъединицам ФСII цианобактерий. Следовательно, две субъединицы РЦ2 пурпурных и две субъединицы ФСII цианобактерий – это результат двух независимых дупликаций генов (рис. 17.11).
Трехмерная структура фотосистем цианобактерий показывает, что они очень близки по пространственной укладке белка и расположению кофакторов – хлорофиллов, феофитинов, хинонов (Baymann, 2001). Однако есть важное различие: в ФСI реакционный центр (в котором происходит разделение зарядов) и антенная часть являются двумя доменами одной белковой цепи, проходящей через мембрану 11 раз, а в ФСII они разделены на отдельные белковые молекулы, кодируемые разными генами. Антенная часть ФСII образуется белками CP43 и CP47, имеющими шесть трансмембранных спиралей, а реакционный центр – белками D1 и D2 c пятью трансмембранными спиралями (рис. 17.12). По трехмерной структуре CP43/CP47 и D1/D2 соответствуют двум доменам единого белка ФСI. РЦ1 Chlorobi и гелиобактерий состоит из двух одинаковых белковых молекул с 11 трансмембранными спиралями каждая, образующими антенный домен и центр разделения зарядов, так же как ФСI цианобактерий. Однако РЦ2 пурпурных бактерий и Chloroflexi не имеют ничего похожего на CP43/CP47 и содержат только пять трансмембранных участков. Функции CP43/CP47 выполняют другие антенные белки, не имеющие никаких аналогов у цианобактерий. По аминокислотной последовательности CP43/CP47 цианобактерий больше похожи на антенный домен РЦ1 гелиобактерий, чем на антенну ФСI той же цианобактериальной клетки. Как нам разобраться, какой вариант фотосистемы древнее – с отдельным антенным белком вроде CP43/CP47 или слитный? Хотя в процессе эволюции происходят как слияния, так и разделения белков, в данном случае гораздо более вероятно разделение предкового двухдоменного белка. Все примитивные варианты фотосистем, состоящие из одинаковых половинок (РЦ1 гелиобактерий и Chloroflexi), состоят из двухдоменных 11-спиральных белков. Появление ФСII в таком случае должно было произойти у предков цианобактерий, а РЦ2 пурпурных бактерий и Chlorobi, видимо, произошли от ФСII древних цианобактерий путем утраты CP43/CP47 и приобрели новые антенные белки (рис. 17.12).
Есть и другой способ узнать, каким способом появилось два типа фотосистем. Если две фотосистемы развивались независимо в разных группах бактерий, то на филогенетических деревьях разных генов фотосинтеза должен быть виден глубокий раздел на ветви обладателей РЦ1 и РЦ2. Среди генов, относящихся к фотосинтезу, наиболее широко распространены ферменты синтеза хлорофилла. Филогенетические деревья этих ферментов (Sousa et al., 2013, Gupta, 2012) показывают, что такого раздела нет (рис. 17.13). Наоборот, Chlorobi с РЦ1 и Chloroflexi с РЦ2 на этих деревьях очень близки друг к другу, а пурпурные бактерии и вовсе попадают внутрь группы цианобактерий. Ферменты пурпурных бактерий специфически сходны с ферментами так называемой ветви С цианобактерий. Члены этой ветви – мелкие одноклеточные цианобактерии океанского пикопланктона (клетки размером менее 2 мкм), такие как Prochlorococcus и Synechococcus. Иначе говоря, история ферментов синтеза хлорофилла, так же как история белков фотосистем, лучше согласуется с появлением обеих фотосистем у предка цианобактерий и горизонтальным переносом их в четыре другие группы фотосинтетических бактерий.
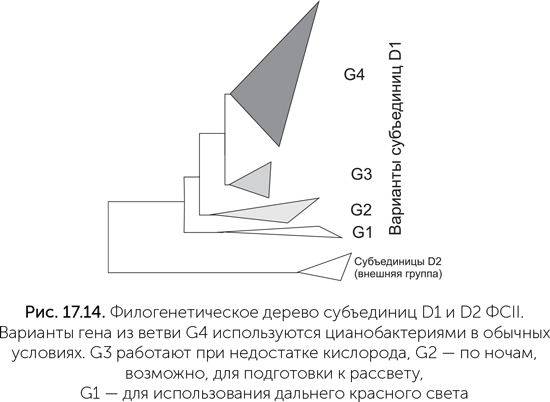
Описанный в главе 16 сценарий появления кислородного фотосинтеза был основан в основном на биофизических экспериментальных данных о современных цианобактериях. Никто тогда не ожидал, что предсказанные переходные формы ФСII, окисляющие марганец и бикарбонат, удастся найти и изучить в пробирке. Однако недавнее исследование геномов цианобактерий показало, что древние варианты ФСII до сих пор существуют и используются клетками в некоторых особых условиях (Cardona, 2015). Водоокисляющий комплекс находится на субъединице D1 – одной из двух неравных половинок реакционного центра ФСII. В геномах многих цианобактерий закодировано несколько (иногда больше десятка) вариантов D1. Было известно, что смена вариантов D1 используется цианобактериями, например, для защиты от слишком яркого света. В работе Танаи Кордона с коллегами было обнаружено, что некоторые варианты субъединицы D1 сильно отличаются от основных, повседневно используемых вариантов. На филогенетическом дереве такие нетипичные белки образуют три отдельные ветви близко к его корню (на рис. 17.14 обозначены G1, G2, G3. Обычные субъединицы D1 образуют ветвь G4). Поскольку у всех известных цианобактерий есть белки из ветви G4 (а они отвечают за окисление воды и выделение кислорода в процессе фотосинтеза), выходит, что белки ветвей G1-G3 появились в результате удвоения генов еще до появления общего предка современных цианобактерий. Ни один из этих белков раньше не попадал в руки экспериментаторов, поэтому об особенностях работы этих белков мы пока можем судить лишь на основе биоинформационного анализа, опираясь на последовательность аминокислот в белках и, соответственно, их структурных свойств и на данные об активности генов в разных условиях.
Последовательности этих необычных вариантов генов D1 показывают, что фотосистемы групп G1 и G2 не имеют аминокислот, необходимых для связывания марганцевого кластера. А значит, они не могут окислять воду. Группа G3 в этом отношении похожа на обычные фотосистемы и, видимо, может окислять воду и выделять кислород. Гены группы G1 включаются при приспособлении цианобактерий к дальнему красному свету (длина волны 720 нм и более, не поглощается обычными фотосистемами), G2 – по ночам, а G3 – при недостатке кислорода. В ближайшие несколько лет биологи-экспериментаторы должны изучить эти цели, разведанные биоинформатиками, и разобраться в деталях работы древних вариантов фотосистемы II. Их результаты должны серьезно уточнить и дополнить описанный путь эволюции кислородного фотосинтеза. |
Последнее изменение этой страницы: 2019-06-19; Просмотров: 71; Нарушение авторского права страницы