|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Дальнейшая эволюция эукариот⇐ ПредыдущаяСтр 79 из 79
После установления симбиоза с митохондриями первые эукариоты стали высокоэффективными хищниками: аэробное дыхание повышает эффективность использования пищи почти в 20 раз по сравнению с гликолизом. Они вышли на поверхность мата и стали поедать цианобактерии. Иногда эукариоты не переваривали съеденные клетки цианобактерий, а оставляли их жить внутри пищеварительной вакуоли и потребляли только выделяемые ими сахара. Такие цианобактерии дали начало второму типу симбиотических органелл – хлоропластам. Эукариоты, приручившие цианобактерий, дали начало трем современным группам водорослей: зеленым, красным и глаукофитовым. Хлоропласты всех этих водорослей покрыты двумя мембранами: внешней – из пищеварительной вакуоли хозяина и внутренней – из клеточной мембраны симбионта. Этот симбиоз называется первичным. Водоросли больше не нуждались в питании фагоцитозом и ушли из бактериального мата в толщу океана, другие же продолжили питаться бактериями. В дальнейшем среди эукариот появились крупные хищные формы, такие как инфузории, приспособившиеся к питанию другими эукариотами, в первую очередь водорослями. Они вместе со своей добычей составили новую экосистему открытого моря, независимую от занявших мелководья бактериальных матов. Эукариоты легко заводят внутриклеточные симбиотические бактерии и подчиняют их себе. Так альфа-протеобактерии стали митохондриями, затем цианобактерии – хлоропластами. И в дальнейшем всякий раз, когда эукариотам требовался какой-нибудь новый биохимический путь, они брали на работу бактерий, уже владеющих им. Так, десятки групп глубоководных животных независимо приручили хемосинтетические бактерии, окисляющие сероводород или метан. Фиксация азота, разложение целлюлозы, синтез витаминов и многое другое эукариоты получают благодаря симбиотическим бактериям. Более того, эукариоты способны вступать в эндосимбиоз с другими эукариотами. Многие группы водорослей возникли в результате вторичного симбиоза, когда в роли хозяев выступали хищные амебы и жгутиконосцы, а симбионтами становились зеленые и красные водоросли (рис. 18.15). При таком симбиозе клетки водорослей дегенерируют, и от них могут остаться только хлоропласт и одна-две дополнительные мембраны. Например, хлоропласты амебы Chlorarachnion происходят от зеленых водорослей. Эти хлоропласты покрыты четырьмя мембранами (две мембраны хлоропласта зеленой водоросли, клеточная мембрана зеленой водоросли и пищеварительная вакуоль амебы). Между второй и третьей мембранами находится нуклеоморф – маленький остаток ядра зеленой водоросли с тремя хромосомами, несущими около 280 генов. Эвгленовые хлоропласты тоже происходят от зеленых водорослей, но дегенерация зашла дальше: нуклеоморфа нет, а из четырех мембран осталось три.
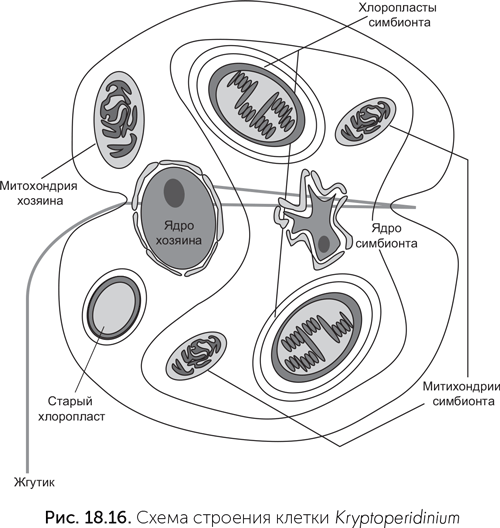
Другой хищный жгутиконосец приручил красную водоросль. Его потомки дали начало бурым, золотистым, диатомовым, криптофитовым, гаптофитовым водорослям и динофлагеллятам. Хлоропласты всех этих водорослей имеют три или четыре мембраны. У криптофитовых хлоропласты содержат нуклеоморф, как у Chlorarachnion. Многие группы динофлагеллятов, даже получив фотосинтезирующих симбионтов, возвращались к хищничеству или переходили к паразитизму. От последних произошли споровики – группа одноклеточных, к которым относится возбудитель малярии. Предки споровиков, как и ряд других водорослей, перенесли в хлоропласт синтез липидов. Поэтому, отказавшись от фотосинтеза, потерять хлоропласт полностью они не смогли. Даже у малярийного плазмодия, предки которого сотни миллионов лет были паразитами, сохраняется апикопласт – маленький, покрытый четырьмя мембранами бесцветный остаток хлоропласта со своей кольцевой ДНК. Среди вернувшихся к хищному образу жизни динофлагеллят есть множество примеров повторного приобретения водорослей-симбионтов. Так, Lepidodinium завел себе зеленую водоросль, Karenia – гаптофитовую, а Dinophysis – криптофитовую (Keeling, 2004). Вершиной же симбиотического таланта эукариот можно считать клетку динофлагелляты Kryptoperidinium. Эта одноклеточная водоросль происходит от динофлагеллят, которые имели хлоропласт – потомок красной водоросли. Затем эти динофлагелляты вернулись к хищному образу жизни. Старый хлоропласт остался у них в качестве маленького фоторецептора (глазка). Потом эти хищные жгутиконосцы вступили в симбиоз с диатомовой водорослью, которая сохранила ядро и значительную часть генома. В клетке Kryptoperidinium под управлением ядра находятся в общей сложности пять «чужих» геномов: свой митохондриальный, старого хлоропласта (фоторецептор), ядерный симбионта-диатомеи, митохондриальный геном симбионта-диатомеи и хлоропластный геном симбионта-диатомеи (рис. 18.16, Figueroa et al., 2009). Деление ядер хозяина и симбионта строго синхронизировано. Более того, при половом размножении происходят мейоз и слияние как главных ядер половых клеток, так и ядер симбионтов.
Наличие цитоскелета и сложной системы регуляции генов позволило эукариотным клеткам объединиться в крупные многоклеточные организмы. Сначала это были нитчатые и лентовидные водоросли, которые ускорили накопление кислорода в атмосфере. Новый уровень кислородного насыщения среды открыл возможность появления многоклеточных животных. За этим последовало радикальное усложнение биосферы («Кембрийский взрыв»), когда за короткое время появились десятки типов животных, и некоторые из ранних представителей быстро достигли метровых размеров. После «Кембрийского взрыва» эволюция шла с ускорением, и с тех пор облик Земли определяют многоклеточные растения и животные.
Заключение: о случайности и закономерности в эволюции, внеземных цивилизациях и о том, зачем Земле люди Одна из загадок, будоражащих умы человечества, – это молчание космоса. Мы до сих пор не нашли никаких достоверных следов других цивилизаций. Если все процессы на пути от неживой материи до цивилизации разумных существ закономерны, то только в нашей Галактике, содержащей примерно 100 млрд звезд, должно существовать множество разумных видов. Если хоть одна цивилизация развивается до строительства межзвездных кораблей, то за небольшое по космическим меркам время – считанные миллионы лет – она может заселить всю Галактику. Но мы не встретили инопланетян и не нашли следов посещения ими Земли в прошлом. В 1960 году шло активное обсуждение программы поиска внеземного разума (SETI). Астроном Фрэнк Дональд Дрейк предложил использовать для оценки количества внеземных цивилизаций, которые мы имеем шанс найти, уравнение, которое впоследствии было названо его именем. Уравнение Дрейка выглядит так: N = R × f p × n e × f l × f i × f c × L, где N – количество разумных цивилизаций, готовых вступить в контакт; R – количество звезд, образующихся в Галактике за год; f p – доля звезд, обладающих планетами; n e – среднее количество планет с подходящими для жизни условиями в планетной системе; f l – вероятность зарождения жизни на планете с подходящими условиями; f i – вероятность развития жизни до разумных форм; f c – вероятность развития разумной жизни до состояния, когда она способна к контакту и ищет его; L – время жизни цивилизации, способной к контакту. Величина R была известна с хорошей точностью уже в 1960 году – в нашей Галактике в среднем возникает семь звезд в год. Два следующих множителя Дрейк оценил в 0, 5 планетных систем на звезду и две планеты, пригодные для жизни, на систему. Остальные четыре величины оценить гораздо сложнее. Если принять, как это сделал Дрейк, вероятность возникновения жизни на подходящей планете в 1, две последние вероятности – в 0, 01, а время жизни цивилизации, способной к контакту, в 10 000 лет, то можно ожидать, что сейчас в нашей Галактике есть 10 цивилизаций, способных к контакту. Исходя из этих оценок развивалась программа SETI, основанная прежде всего на анализе сигналов, принимаемых радиотелескопами. Сейчас мы можем точнее оценить многие множители уравнения Дрейка. Тысячи открытых экзопланет подтверждают старую оценку fp = 0, 5. Однако подавляющее большинство этих планет находится слишком близко к звезде, и они слишком горячи, чтобы быть обитаемыми. Поэтому современные оценки ne более пессимистичны, чем предполагали в 1960-е годы, и лежат в интервале 0, 001–0, 1 (см. главу 2). Ближе к теме основной части книги относятся следующие два множителя – вероятность возникновения жизни на планете, пригодной для этого, и вероятность развития возникшей жизни до разумных форм. Вероятность возникновения жизни (fl) оценить сложно по двум причинам. Во-первых, мы знаем только об одном таком событии. По одному событию нельзя оценить его вероятность. Во-вторых, трудно определить, какие планеты считать «подходящими». Например, в современной Солнечной системе Марс и Европа подходят для жизни некоторых земных микробов, но жизнь земного типа там сейчас возникнуть не может. Дрейк взял вероятность возникновения жизни на «подходящей» планете за единицу, т. е. имел в виду «подходящую для возникновения жизни» планету, а не «подходящую для существования». Долю планет, подходящих для существования жизни земного типа, астрономы сейчас оценивают в пределах от 0, 1 до 0, 001. Доля подходящих для возникновения жизни планет явно меньше, и неизвестно, насколько. Поэтому большие надежды возлагаются на исследования Марса. Это единственное кроме Земли место в Солнечной системе, где теоретически могла появиться жизнь земного типа (водно-углеродная с ДНК и белками). Если следы возникновения жизни на Марсе будут найдены, значит, в Солнечной системе две планеты подходили для возникновения жизни, а значит, и в Галактике их много. Если же Земля уникальна в Солнечной системе, то трудно оценить, насколько она уникальна в Галактике. Современные методы поиска экзопланет могут показать, что некая планета похожа на Землю по массе, температуре и наличию воды. Но она может быть, например, полностью покрыта океаном стокилометровой глубины, а для возникновения жизни, основанной на белках и нуклеиновых кислотах, нужны пересыхающие лужи. Мы не можем определить, есть ли у экзопланеты спутники, а Луна явно внесла вклад в появление жизни на Земле. Короче, мы можем считать, что вероятность возникновения жизни на планете, очень похожей на древнюю Землю, достаточно велика (скажем, 0, 5). Но мы не знаем, насколько часто такие планеты встречаются в Галактике. Ведь важны сразу многие параметры: масса, расстояние от звезды, история с мегаимпактом, спутник, содержание воды, состав атмосферы, осевое вращение, стабильность орбиты и минимум астероидных ударов. И мы не знаем, насколько большие отклонения от земных параметров допустимы для появления жизни. О следующем множителе уравнения – fi, вероятности развития жизни до разумных существ, у нас больше информации. Эволюция от первых форм жизни до высших животных состоит из множества шагов. Отдельные эволюционные шаги не уникальны, их можно сравнивать друг с другом и оценивать вероятность. И здесь мы сталкиваемся с серьезным недостатком уравнения Дрейка: оно не учитывает времени, необходимого на возникновение и развитие жизни. Это время, судя по примеру земной жизни, весьма велико: от первых достоверных следов жизни до появления человеческой цивилизации прошло 3, 5 млрд лет – четверть возраста Вселенной! Есть основания считать, что возраст земной жизни даже больше 3, 5 млрд лет и может достигать 4, 2–4, 3 млрд лет. Поэтому пренебрегать этим сроком нельзя даже в масштабе возраста Вселенной. Может быть, небольшие различия начальных условий на планете могут удлинить этот срок в полтора-два раза и люди просто возникли раньше других разумных видов? Попробуем разобраться, чем определяются сроки различных этапов эволюции жизни. Напомним основные даты истории жизни на нашей планете: • 4, 3 млрд лет назад – остывание планеты после мегаимпакта. Самый ранний срок, когда жизнь теоретически могла возникнуть; • 3, 5 млрд лет назад – первые следы сложных микробных сообществ, состоящих из клеток, похожих на современные бактерии; • 2, 4 млрд лет назад – появление кислородного фотосинтеза, первое глобальное вымирание, появление эукариот; • между 800 и 600 млн лет назад – появление многоклеточных животных. Мы не знаем точно, когда появились первые формы жизни на нашей планете. Но можно с достаточной уверенностью судить, что они населяли только определенное местообитание – наземные геотермальные водоемы (глава 6). Лишь с возникновением мембран, не пропускающих ионы (глава 15), клетки смогли выйти за пределы геотермальной колыбели. До этого примитивная жизнь была уязвима к колебаниям климата и геологической активности родного геотермального поля. Все промежуточные стадии на этом пути (главы 9–15) были поддержаны естественным отбором, и нет никаких причин, по которым переход от РНК-мира к клеткам мог бы затянуться. Следовательно, путь от РНК-мира до клеток бактерий и архей был пройден достаточно быстро, в пределах 10–100 млн лет – любая задержка привела бы к вымиранию. Затем эволюция жизни замедлилась. Более 1 млрд лет (промежуток от 3, 5 до 2, 4 млрд лет назад) никаких крупных эволюционных событий не происходило. Лишь появление кислородного фотосинтеза 2, 4 млрд лет назад резко подстегнуло эволюцию. Кислородное отравление уничтожило большинство древних видов микробов, освободившиеся экологические ниши стали занимать молодые группы, такие как альфа-протеобактерии и археи Sulfolobales. Что определило задержку эволюции на этапе бескислородной микробной жизни на миллиард с лишним лет? Как мы видели в главе 16, кислородный фотосинтез возник на основе марганец-окисляющего фотосинтеза. Следы осаждения окисленного марганца фотосинтезирующими организмами есть в морских осадках возраста 2, 4 млрд лет. Другие известные варианты бескислородного фотосинтеза используют более распространенные и легче окисляемые вещества – серу, сероводород и соединения железа. Следы железоокисляющего фотосинтеза – полосатые железистые отложения (джеспилиты) обычны в морских осадках возраста 3, 5–2, 4 млрд лет, а потом их образование прекратилось. Переход к окислению марганца, а затем к кислородному фотосинтезу, скорее всего, был вызван исчерпанием запасов растворенных солей железа в океане. Пока железа хватало, возникновение более сложных форм фотосинтеза не поддерживалось отбором. Баланс растворенного железа в древних морях складывался из прихода (выброс железа «черными курильщиками» и смыв с суши) и расхода (осаждение фотосинтезирующими клетками и медленное абиогенное осаждение). Все поступление железа в море в конечном итоге связано с геологической активностью планеты и содержанием железа в ее верхних слоях. В ходе эволюции планеты железо из мантии постепенно переходит в ядро. Современная верхняя мантия Земли, из которой образуется новая океанская кора, заметно беднее железом, чем 3, 5 млрд лет назад. Темпы осаждения железа к ядру планеты зависят от ее размера: чем больше планета, тем дольше идет процесс. Следовательно, на массивной планете (суперземле) «кислородная революция» и все дальнейшие этапы эволюции жизни будут отложены, возможно, на миллиарды лет. Другие факторы, которые могут оттянуть «кислородную революцию», – это любые ограничения продуктивности морских фотосинтезирующих микробов. Недостаток фосфора, растворимых форм азота, оледенение заметной части планеты – все эти факторы уменьшают продуктивность фотосинтеза и темпы осаждения железа. Кислородная революция, судя по всему, стала толчком для появления нового уровня сложности – эукариотной клетки. Только из эукариотных клеток, обладающих цитоскелетом и сложной регуляцией работы генов, можно построить многоклеточный организм из разных типов клеток с разделением труда между ними. Бактерии и археи к этому не способны. Как мы видели в главе 18, появление эукариот включало в себя череду событий, которые сложно назвать закономерными. Сначала архея-метаноген, пострадав от кислорода, смогла перейти к совершенно другому обмену веществ – брожению. Потом она вступила в симбиоз с предками митохондрий. Прецедентов такого симбиоза среди бактерий и архей неизвестно! Возможно, в симбиозе принял участие и крупный ДНК-вирус. Все эти шаги были пройдены не благодаря естественному отбору, а, скорее, вопреки ему. Роль отбора и других эволюционных процессов, таких как дрейф генов, в появлении эукариот подробно рассматривал Е. Кунин в книге «Логика случая». На современном уровне знаний создается впечатление, что появление эукариот не было закономерно, в отличие от появления клеток. Вероятность развития жизни до эукариотного уровня оценить сложно, но она может быть достаточно мала. Вполне возможно, что миллионы планет в нашей Галактике заселены жизнью бактериального уровня сложности, но только на нескольких возникло что-то сравнимое с эукариотами. Следующий ключевой этап на пути к разуму – это многоклеточные организмы. Многоклеточность среди эукариот возникала много раз: животные, настоящие грибы, оомицеты, три независимо возникших линии слизевиков, наземные растения, бурые водоросли, красные водоросли. Так что этот этап можно считать закономерным и неизбежным, но остается вопрос о сроках. От кислородной революции (и, видимо, появления эукариот) до достижения многоклеточного уровня прошло 1, 5 млрд лет. Первые несомненные остатки животных имеют возраст 635 млн лет (эдиакарская фауна, или вендобионты). Эти животные не имели рта, конечностей и скелета, об их способе питания и родстве с современными группами животных ученые спорят. Остатки современных типов животных, обладающих скелетом (членистоногие, моллюски, губки), появляются 540 млн лет назад. Этот момент истории Земли называется «Кембрийским взрывом». Тогда произошло лавинообразное возрастание разнообразия и сложности животных, появление разных типов скелетов, распространение хищников. Молекулярные часы указывают на более раннее появление животных, около 700–800 млн лет назад, но первые животные, видимо, были микроскопическими и бесскелетными и не сохранились в ископаемом состоянии. Зато сохранились окаменевшие одноклеточные водоросли, среди которых в период 750–700 млн лет назад происходили резкие изменения размеров и появление шипастых клеток. Это могло быть ответом на выедание первыми животными. Для возникновения многоклеточных животных на Земле было необходимо накопление заметного количества кислорода в атмосфере и океане (подробно изложено у К. Еськова в «Истории Земли и жизни на ней»). Накопление кислорода началось 2, 4 млрд лет назад, но шло медленно. Лишь примерно 800 млн лет назад концентрация кислорода стала лавинообразно нарастать и к моменту «Кембрийского взрыва» 540 млн лет назад превысила половину от современной. Подобно железу, баланс кислорода складывается из прихода и расхода. Приход полностью определяется продуктивностью фотосинтеза, а расход зависит от множества факторов. Кислород расходуется на окисление органики и материковых горных пород. Захоронение неокисленных останков живых организмов приводит к накоплению кислорода в атмосфере, а эти останки превращаются в сланцы и нефть. На баланс между окислением и захоронением органических веществ влияет множество причин, включая соотношение суши и моря, площадь морей разной глубины, структуру морских течений и геологическую активность. Расход кислорода на окисление минералов также определяется геологической активностью, выносящей на поверхность окисляемые минералы, такие как базальт и пирит. Продуктивность фотосинтеза может ограничиваться холодами, недостатком фосфора или света. Недостаток света был вызван мутностью морской воды: до появления наземных растений вся суша сильно размывалась дождями, песок и глина сносились в моря, и мелкая глинистая взвесь делала весь океан мутным. Геологические данные показывают, что моря стали прозрачными и более продуктивными во время «Кембрийского взрыва», и очистили их животные-фильтраторы. В целом можно сказать, что сроки появления многоклеточных животных, судя по всему, велики и легко могли сдвинуться на миллиард-другой лет позже по самым разным причинам. Многоклеточные животные и растения, особенно вышедшие на сушу, более уязвимы к глобальным катастрофам, чем микробная биосфера. Микробы пережили (хотя и с потерями) позднюю метеоритную бомбардировку, а подземная микробная биосфера вообще практически неуязвима. Эволюцию животных и растений может оборвать или отбросить на сотни миллионов лет назад один астероид или глобальное оледенение («Земля-снежок»). Таких глобальных оледенений на Земле было два: первое – 2, 3 млрд лет назад, сразу за кислородной революцией, другое – 800–650 млн лет назад, примерно во времена появления первых животных. О причинах этих глобальных оледенений ученые спорят. Есть и другие варианты катастрофы, приводящей к глобальному вымиранию многоклеточной жизни, например близкая вспышка сверхновой. Орбита Солнца в Галактике проходит вдали от районов частых вспышек сверхновых, но это не дает полной гарантии безопасности. Многие другие звезды расположены менее благоприятно в этом смысле, и жизнь на поверхности их планет может гибнуть от вспышек сверхновых каждые 200–500 млн лет. Относительную стабильность климата Земли обеспечивает Луна. Взаимодействие с Луной ограничивает колебания оси вращения Земли и, следовательно, колебания климата. Марс, не имеющий такого спутника, испытывает колебания наклона оси вращения от 0 до 40 градусов и частые изменения климата. Без Луны такие колебания климата на Земле, скорее всего, приводили бы к частым вымираниям животных и оттянули бы появление разумного вида. Итак, мы видим, что в случае жизни земного типа множитель fi (вероятность развития жизни до разумных форм) может быть очень мал. Причин этому минимум три: возможная задержка с появлением кислородного фотосинтеза, сомнительная вероятность появления эукариот и задержка эволюции многоклеточности. Со всеми этими поправками к уравнению Дрейка вполне возможно, что за всю историю нашей Галактики в ней возникли считанные десятки, а то и единицы разумных видов. Если что-то помешало им начать космическую экспансию, скорее всего, каждый из них вымер еще до появления другого, и ни один из видов не имел шанса вступить в контакт. Исследования, которые проясняют новые детали происхождения жизни, как правило, имеют и важное практическое применение. Например, создание и изучение ксенонуклеиновых и пептидонуклеиновых кислот (глава 12) ведется прежде всего для нужд медицины. Очень перспективным считается создание лекарств, действующих по принципу РНК-интерференции. Такие лекарства могут выключать один строго определенный ген в определенном типе клеток, не воздействуя на другие процессы. Их можно быстро синтезировать «под заказ», создавая, например, противораковые лекарства для каждого пациента индивидуально, с учетом особенностей его опухоли. С одной стороны, химическая основа таких лекарств должна быть подобна РНК, чтобы образовывать двойную спираль с матричной РНК выключаемого гена, а с другой – она должна отличаться от РНК, чтобы не быть разрушенной клеточной системой противовирусной защиты. Ксенонуклеиновые кислоты удовлетворяют этим требованиям и могут стать основой нового класса лекарств. Расширение нуклеотидного алфавита (глава 6) и набора аминокислот (глава 13) является важным направлением синтетической биологии. Это позволяет получать линии микробов с новыми, невозможными в рамках 20 аминокислот свойствами, что очень пригодится в биотехнологии. Переработка всевозможных отходов, обезвреживание разливов нефти, биотопливо, новые материалы и многое другое станет доступнее. Изучение биоэнергетики – комплексов дыхательных цепей, фотосистем и белковых проводов (главы 15, 16, 17) – может помочь в решении энергетических проблем человечества. Знание тонкостей работы фотосистем помогает совершенствовать искусственные солнечные батареи. Есть и более смелые исследования: как оказалось, дыхательные цепи бактериальных клеток можно подключить к внешним электродам, и такие «микробные батареи» дают ток. Современные солнечные батареи требуют больших затрат энергии на свое производство, и возможно, их станут вытеснять пластиковые баки с генно-модифицированными цианобактериями. Их клетки могут быть напрямую подключены к проводам или же настроены на производство любого необходимого горючего – водорода, спирта, биогаза или дизельного топлива. В отличие от солнечных батарей, они растут сами. При серьезной генной модификации организмов необходимо принять меры, чтобы искусственные гены не передались другим видам. Здесь может помочь расширение нуклеотидного алфавита – если новые гены будут записаны новыми буквами, дикие микробы не смогут их прочитать. Но самый радикальный способ остановить утечку генов – это создать «зазеркальные» клетки из правых аминокислот и левых нуклеотидов. В природе нет механизмов, которые могли бы перенести информацию из левой ДНК в стандартную правую. Такая «зазеркальная» искусственная жизнь будет полностью подконтрольна людям, а заодно и защищена от всех природных вирусов. Эти перспективы могут показаться пугающими. Не приведут ли последствия таких модификаций к гибели существующих видов? Нет, не приведут. Все природные организмы оптимизированы на выживание и размножение. Творения биоинженеров же должны прежде всего выполнять свою функцию. А живой организм не может быть хорош сразу во всем. За выдающиеся достижения в любой одной области неизбежно приходится платить слабостью в других. Поэтому у модифицированных и полностью искусственных микробов мало шансов в конкуренции с дикими видами. Даже обычные домашние животные, которые не подвергались генным модификациям, успешно дичают только в отсутствие конкурентов и хищников, например на островах. Организмы со значительными генными модификациями будут еще более несбалансированными и тем более уступят диким конкурентам. «Зазеркальный» микроб к тому же будет отрезан от обмена генами с другими организмами, что окончательно подорвет его эволюционные перспективы. Вообще, противостояние человека и природы, рисуемое экологической пропагандой, несколько преувеличено. Внутреннее событие биосферы – «кислородная революция» цианобактерий – вызвало более масштабное вымирание, чем все падения астероидов, глобальные оледенения и тем более деятельность человечества. Даже глобальная ядерная война будет иметь меньшие последствия, чем появление кислородного фотосинтеза. Кстати, в зоне отчуждения вокруг Чернобыльской АЭС уже через несколько лет после катастрофы фактически образовался заповедник, населенный большим количеством оленей, кабанов, волков, лис и других животных. Радиоактивное заражение мешает им гораздо меньше, чем вырубка лесов и сельское хозяйство. Какие действия человека считаются вредными для биосферы? Например, люди добывают нефть, газ и уголь, чтобы сжечь их. Но это не чужое для биосферы вещество. Это остатки древних растений и водорослей, это углерод, который захоронился и выпал из глобального круговорота в древние эпохи. Человек просто возвращает биосфере потерянное ею. Глобальное потепление и таяние ледников, которое может произойти из-за сжигания нефти, – это всего лишь возвращение Земли к нормальному для нее климату. Оледенения на полюсах за последние полмиллиарда лет были лишь два раза: в конце каменноугольного и пермском периодах (310–250 млн лет назад) и последние 40 млн лет. В остальные эпохи на большей части суши до самого полюса климат был близок к субтропическому. Например, ископаемые остатки крокодилов возраста 50 млн лет найдены на острове Элсмир (Канадский арктический архипелаг, далеко за полярным кругом). Так что таяние ледников будет неприятностью лишь для людей. Для Земли это скорее возвращение к норме из длительного ледникового периода. Точно так же люди добывают фосфориты и производят из них удобрения для полей. Так в биосферу возвращается потерянный ею когда-то фосфор. Без человека круговорот фосфора практически не замыкается: фосфор постепенно смывается с суши в море и откладывается в морских осадках. Движение плит океанской коры может занести эти осадки в мантию, откуда фосфор выйдет с вулканическими газами и пеплом и вернется в биосферу. Но на современной Земле тектоническая активность не покрывает осаждения фосфора в океанах. Потеря фосфора постепенно снижает продуктивность всей биосферы. Только человек, добывая похороненные в толще земной коры фосфориты, решает эту проблему биосферы. Есть и еще более важная услуга, которую только разумный вид может оказать родной биосфере. Время, отведенное на развитие жизни на планете, ограничено. Светимость Солнца неуклонно возрастает, и через 1, 5 млрд лет даже полное изъятие углекислого газа из атмосферы не сможет предотвратить глобальное потепление. После этого Земля неизбежно превратится в подобие Венеры, а биосфера погибнет. Иначе говоря, земная жизнь уже потратила три четверти отведенного ей времени. И она неизбежно погибнет, если разумные существа не построят космические корабли и не возьмут земные организмы с собой к другим звездам. В этом смысле технологическое развитие человечества необходимо для всего живого на нашей планете.
Библиография Диброва Д., Гальперин М., Кунин Е., Мулкиджанян А. Древние системы натрий-калиевого гомеостаза клетки как предшественники мембранной биоэнергетики // Биохимия, 2015. Т. 80. Вып. 5. С. 590–611. Еськов К. Удивительная палеонтология: История Земли и жизни на ней. М., 2007. Кунин Е. Логика случая. О природе и происхождении биологической эволюции. М., 2014. Марков А., Куликов А. Гомологичные белковые домены в надцарствах Archaea, Bacteria и Eukaryota и проблема происхождения эвкариот // Известия Российской академии наук. Серия биологическая. 2005. № 4. 389–400. Марков А., Куликов А. Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы // Палеонтологический журнал. 2005. № 4. С. 3–18. Марков А., Куликов А. Происхождение эукариот как результат интеграционных процессов в микробном сообществе (http: //evolbiol.ru/dok_ibr2009.htm) Acevedo O. L. & Orgel L. E. Non-enzymatic transcription of an oligodeoxynucleotide 14 residues long // Journal of Molecular Biology, 1987, vol. 197, pp. 187–193. DOI: 10.1016/0022–2836(87)90117–3 Adamala K. & Szostak J. W. Competition between model protocells driven by an encapsulated catalyst // Nature Chemistry, 2013, vol. 5, pp. 495–501. DOI: 10.1038/nchem.1650 Adamala K. et al. Open questions in origin of life: experimental studies on the origin of nucleic acids and proteins with specific and functional sequences by a chemical synthetic biology approach // Computational and Structural Biotechnology Journal, 2014, vol. 9. DOI: 10.5936/csbj.201402004 Attwater J., Wochner A. & Holliger P. In-ice evolution of RNA polymerase ribozyme activity // Nature Chemistry, 2013, vol. 5, pp. 1011–1018. DOI: 10.1038/nchem.1781 Baaske P. et al. Extreme accumulation of nucleotides in simulated hydrothermal pore systems // Proceedings of the National Academy of Sciences, 2007, vol. 104, pp. 9346–9351. DOI: 10.1073/pnas.0609592104 Bailey S., Wing R. A. & Steitz T. A. The Structure of T. aquaticus DNA Polymerase III Is Distinct from Eukaryotic Replicative DNA Polymerases // Cell, 2006, vol. 126, pp. 893–904. DOI: 10.1016/j.cell.2006.07.027 Bali S. et al. Molecular hijacking of siroheme for the synthesis of heme and d1 heme // Proceedings of the National Academy of Sciences, 2011, vol. 108, pp. 18260–18265. DOI: 10.1073/pnas.1108228108 Bar-Even A., Noor E., Flamholz A. & Milo R. Design and analysis of metabolic pathways supporting formatotrophic growth for electricity-dependent cultivation of microbes // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2013, vol. 1827, pp. 1039–1047. DOI: 10.1016/j. bbabio.2012.10.013 Barry R. D. The multiplication of influenza virus: II. Multiplicity reactivation of ultraviolet irradiated virus // Virology, 1961, vol. 14, pp. 398–405. DOI: 10.1016/0042–6822(61)90330–0 Baum D. A. & Baum B. An inside-out origin for the eukaryotic cell // BMC Biology, 2014, vol. 12, p. 76. DOI: 10.1186/s12915-014-0076-2 Baymann, F. et al. The redox protein construction kit: pre-last universal common ancestor evolution of energy-conserving enzymes // Philosophical Transactions of the Royal Society of London B: Biological Sciences, 2003, vol. 358, pp. 267–274. DOI: 10.1098/rstb.2002.1184 Baymann, F., Brugna, M., Mü hlenhoff, U. & Nitschke, W. Daddy, where did (PS) I come from? // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2001, vol. 1507, pp. 291–310. DOI: 10.1016/S0005–2728(01)00209–2 Bell, P. J. L. The viral eukaryogenesis hypothesis: a key role for viruses in the emergence of eukaryotes from a prokaryotic world environment // Annals of the New York Academy of Sciences, 2009, vol. 1178, 91–105. DOI: 10.1111/j.1749–6632.2009.04994.x Bell, P. J. Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus? // Journal of molecular evolution, 2001, vol. 53, pp. 251–256. DOI: 10.1007/s002390010215 Bell, P. J. L. Sex and the eukaryotic cell cycle is consistent with a viral ancestry for the eukaryotic nucleus // Journal of theoretical biology, 2006, vol. 243, pp. 54–63. DOI: 10.1016/j. jtbi.2006.05.015 Benner, S. A., Kim, H.-J., Kim, M.-J. & Ricardo, A. Planetary Organic Chemistry and the Origins of Biomolecules // Cold Spring Harbor Perspectives in Biology, 2010, vol. 2, p. a003467. DOI: 10.1101/cshperspect.a003467 Benner, S. A., Kim, H.-J. & Carrigan, M. A. Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA // Accounts of Chemical Research, 2012, vol. 45, pp. 2025–2034. DOI: 10.1021/ar200332w Benner, S. A., Ricardo, A. & Carrigan, M. A. Is there a common chemical model for life in the universe? // Current Opinion in Chemical Biology, 2004, vol. 8, pp. 672–689. DOI: 10.1016/j. cbpa.2004.10.003 Bernhardt, H. S. & Sandwick, R. K. Purine Biosynthetic Intermediate-Containing Ribose-Phosphate Polymers as Evolutionary Precursors to RNA // Journal of Molecular Evolution, 2014, vol. 1–14. DOI: 10.1007/s00239-014-9640-1 Bokov, K. & Steinberg, S. V. A hierarchical model for evolution of 23S ribosomal RNA // Nature, 2009, vol. 457, pp. 977–980. DOI: 10.1038/nature07749 Borowska, Z. & Mauzerall, D. Photoreduction of carbon dioxide by aqueous ferrous ion: An alternative to the strongly reducing atmosphere for the chemical origin of life // Proceedings of the National Academy of Sciences, 1988, vol. 85, pp. 6577–6580. Bortnikova, S. B., Gavrilenko, G. M., Bessonova, E. P. & Lapukhov, A. S. The hydrogeochemistry of thermal springs on Mutnovskii Volcano, southern Kamchatka // Journal of Volcanology and Seismology, 2010, vol. 3, pp. 388–404. DOI: 10.1134/S0742046309060025 Braakman, R. & Smith, E. The Emergence and Early Evolution of Biological Carbon-Fixation // PLoS Computational Biology, 2012, vol. 8. DOI: 10.1371/journal.pcbi.1002455 Breslow, R. & Cheng, Z.-L. L-amino acids catalyze the formation of an excess of D-glyceraldehyde, and thus of other D sugars, under credible prebiotic conditions // Proceedings of the National Academy of Sciences, 2010, vol. 107, pp. 5723–5725. DOI: 10.1073/pnas.1001639107 Breslow, R. & Levine, M. S. Amplification of enantiomeric concentrations under credible prebiotic conditions // Proceedings of the National Academy of Sciences, 2006, vol. 103, pp. 12979–12980. DOI: 10.1073/pnas.0605863103 Breslow, R. The origin of homochirality in amino acids and sugars on prebiotic earth // Tetrahedron Letters, 2011, vol. 52, pp. 4228–4232. DOI: 10.1016/j. tetlet.2011.06.002 Briones, C., Stich, M. & Manrubia, S. C. The dawn of the RNA World: Toward functional complexity through ligation of random RNA oligomers // RNA, 2009, vol. 15, pp. 743–749. DOI: 10.1261/rna.1488609 Buchanan, B. B. & Arnon, D. I. A reverse KREBS cycle in photosynthesis: consensus at last // Photosynthesis Research, 1990, vol. 24, pp. 47–53. Budisa, N. & Schulze-Makuch, D. Supercritical Carbon Dioxide and Its Potential as a Life-Sustaining Solvent in a Planetary Environment // Life, 2014, vol. 4, pp. 331–340. DOI: 10.3390/life4030331 Cahová, H., Winz, M.-L., Hö fer, K., Nü bel, G. & Jä schke, A. NAD captureSeq indicates NAD as a bacterial cap for a subset of regulatory RNAs // Nature advance online publication, 2014. DOI: 10.1038/nature14020 Canup, R. M. & Asphaug, E. Origin of the Moon in a giant impact near the end of the Earth's formation // Nature, 2001, vol. 412, pp. 708–712. DOI: 10.1038/35089010 Canup, R. M. & Ward, W. R. A common mass scaling for satellite systems of gaseous planets // Nature, 2006, vol. 441, pp. 834–839. DOI: 10.1038/nature04860 Canup, R. M. Origin of Saturn/'s rings and inner moons by mass removal from a lost Titan-sized satellite // Nature, 2010, vol. 468, pp. 943–946. DOI: 10.1038/nature09661 Cardona, T., Murray, J. W. & Rutherford, A. W. Origin and Evolution of Water Oxidation before the Last Common Ancestor of the Cyanobacteria // Molecular Biology and Evolution, 2015, vol. 32, pp. 1310–1328 (2015). DOI: 10.1093/molbev/msv024 Carter Jr., C. W. & Duax, W. L. Did tRNA Synthetase Classes Arise on Opposite Strands of the Same Gene? // Molecular Cell, 2002, vol. 10, pp. 705–708 (2002). DOI: 10.1016/S1097–2765(02)00688–3 Carter, C. W. et al. The Rodin-Ohno hypothesis that two enzyme superfamilies descended from one ancestral gene: an unlikely scenario for the origins of translation that will not be dismissed // Biology Direct, 2014, vol. 9, pp. 11. DOI: 10.1186/1745-6150-9-11 Cernak, P. & Sen, D. A thiamin-utilizing ribozyme decarboxylates a pyruvate-like substrate. Nature Chemistry, 2013, vol. 5, pp. 971–977. DOI: 10.1038/nchem.1777 Chen, J. & Strous, M. Denitrification and aerobic respiration, hybrid electron transport chains and co-evolution // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2013, vol. 1827, pp. 136–144. DOI: 10.1016/j. bbabio.2012.10.002 Chen, X., Li, N. & Ellington, A. D. Ribozyme Catalysis of Metabolism in the RNA World // Chemistry & Biodiversity, 2007, vol. 4, pp. 633–655. DOI: 10.1002/cbdv.200790055 Chinnapen, D. J.-F. & Sen, D. A deoxyribozyme that harnesses light to repair thymine dimers in DNA // Proceedings of the National Academy of Sciences of the United States of America, 2004, vol. 101, pp. 65–69. DOI: 10.1073/pnas.0305943101 Chrysostomou, A. et al. Polarimetry of young stellar objects – III. Circular polarimetry of OMC-1 // Monthly Notices of the Royal Astronomical Society, 2000, vol. 312, pp. 103–115. DOI: 10.1046/j. 1365–8711.2000.03126.x Cleaves II, H. J. The prebiotic geochemistry of formaldehyde // Precambrian Research, 2008, vol. 164, pp. 111–118. DOI: 10.1016/j. precamres. 2008.04.002 Cleaves, H. J. & Miller, S. L. The Nicotinamide Biosynthetic Pathway Is a By-Product of the RNA World // Journal of Molecular Evolution, 2001, vol. 52, pp. 73–77. DOI: 10.1007/s002390010135 Connerney, J. E. P. et al. Tectonic implications of Mars crustal magnetism // Proceedings of the National Academy of Sciences of the United States of America, 2005, vol. 102, pp. 14970–14975. DOI: 10.1073/pnas.0507469102 Copley, S. D., Smith, E. & Morowitz, H. J. A mechanism for the association of amino acids with their codons and the origin of the genetic code // Proceedings of the National Academy of Sciences of the United States of America, 2005, vol. 102, pp. 4442–4447. DOI: 10.1073/pnas.0501049102 Costanzo, G., Pino, S., Ciciriello, F. & Mauro, E. D. Generation of Long RNA Chains in Water // Journal of Biological Chemistry, 2009, vol. 284, pp. 33206–33216. DOI: 10.1074/jbc.M109.041905 Costanzo, G., Saladino, R., Crestini, C., Ciciriello, F. & Mauro, E. D. Formamide as the main building block in the origin of nucleic acids // BMC Evolutionary Biology, 2007, vol. 7, S1. DOI: 10.1186/1471-2148-7-S2-S1 Cozens, C., Pinheiro, V. B., Vaisman, A., Woodgate, R. & Holliger, P. A short adaptive path from DNA to RNA polymerases // Proceedings of the National Academy of Sciences, 2012, vol. 109, pp. 8067–8072. DOI: 10.1073/pnas.1120964109 Crida, A. Solar System formation // arXiv: 0903.3008 [astro-ph], 2009. Č uboň ová, L. et al. Archaeal DNA Polymerase D but Not DNA Polymerase B Is Required for Genome Replication in Thermococcus kodakarensis // Journal of Bacteriology, 2013, vol. 195, pp. 2322–2328. DOI: 10.1128/JB.02037–12 Dadachova, E. et al. Ionizing Radiation Changes the Electronic Properties of Melanin and Enhances the Growth of Melanized Fungi // PLoS ONE, 2007, vol. 2. DOI: 10.1371/journal. pone.0000457 Dailey, H. A. Illuminating the black box of B12 biosynthesis // Proceedings of the National Academy of Sciences, 2013, vol. 110, pp. 14823–14824 (2013). DOI: 10.1073/pnas.1313998110 Dailey, H. A., Gerdes, S., Dailey, T. A., Burch, J. S. & Phillips, J. D. Noncanonical coproporphyrin-dependent bacterial heme biosynthesis pathway that does not use protoporphyrin // Proceedings of the National Academy of Sciences, 2015, vol. 112, pp. 2210–2215. DOI: 10.1073/pnas.1416285112 Dawkins. The River out of Eden. Harper Collins, 1996. Desmond, E. & Gribaldo, S. Phylogenomics of Sterol Synthesis: Insights into the Origin, Evolution, and Diversity of a Key Eukaryotic Feature // Genome Biology and Evolution, 2009, vol. 1, pp. 364–381. DOI: 10.1093/gbe/evp036 Di Giulio, M. An extension of the coevolution theory of the origin of the genetic code // Biology Direct, 2008, vol. 3, pp. 37–37. DOI: 10.1186/1745-6150-3-37 Dibrova, D. V., Cherepanov, D. A., Galperin, M. Y., Skulachev, V. P. & Mulkidjanian, A. Y. Evolution of cytochrome bc complexes: From membrane-anchored dehydrogenases of ancient bacteria to triggers of apoptosis in vertebrates // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2013, vol. 1827, pp. 1407–1427. DOI: 10.1016/j. bbabio.2013.07.006 Dibrova, D. V., Makarova, K. S., Galperin, M. Y., Koonin, E. V. & Mulkidjanian, A. Y. Comparative analysis of lipid biosynthesis in archaea, bacteria and eukaryotes: What was the structure of the first membrane lipids? // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2012, vol. 1817, Supplement, S154. DOI: 10.1016/j. bbabio.2012.06.404 Dismukes, G. C. et al. The origin of atmospheric oxygen on Earth: The innovation of oxygenic photosynthesis // Proceedings of the National Academy of Sciences, 2001, vol. 98, pp. 2170–2175. DOI: 10.1073/pnas.061514798 Douglas, R. H. et al. Dragon fish see using chlorophyll // Nature, 1998, vol. 393, pp. 423–424. DOI: 10.1038/30871 Ducluzeau, A.-L. et al. Was nitric oxide the first deep electron sink? // Trends in Biochemical Sciences, 2009, vol. 34, pp. 9–15. DOI: 10.1016/j. tibs.2008.10.005 Engelhart, A. E., Powner, M. W. & Szostak, J. W. Functional RNAs exhibit tolerance for non-heritable 2`-5` vs. 3`-5` backbone heterogeneity // Nature chemistry, 2013, vol. 5, pp. 390–394. DOI: 10.1038/nchem.1623 Eschenmoser, A. On a Hypothetical Generational Relationship between HCN and Constituents of the Reductive Citric Acid Cycle // Chemistry & Biodiversity, 2007, vol. 4, pp. 554–573. DOI: 10.1002/cbdv.200790050 Ettema, T. J. G., Lindå s, A.-C. & Bernander, R. An actin-based cytoskeleton in archaea // Molecular microbiology, 2011, vol. 80, pp. 1052–1061. DOI: 10.1111/j. 1365–2958.2011.07635.x Fairé n, A. G., Ferná ndez-Remolar, D., Dohm, J. M., Baker, V. R. & Amils, R. Inhibition of carbonate synthesis in acidic oceans on early Mars // Nature, 2004, vol. 431, pp. 423–426. DOI: 10.1038/nature02911 Figueroa, R. I., Bravo, I., Fraga, S., Garcé s, E. & Llaveria, G. The Life History and Cell Cycle of Kryptoperidinium foliaceum, A Dinoflagellate with Two Eukaryotic Nuclei // Protist, 2009, vol. 160, pp. 285–300. DOI: 10.1016/j.protis.2008.12.003 Fisher, M. A., McKinley, K. L., Bradley, L. H., Viola, S. R. & Hecht, M. H. De Novo Designed Proteins from a Library of Artificial Sequences Function in Escherichia Coli and Enable Cell Growth // PLoS ONE, 2011, vol. 6. DOI: 10.1371/journal.pone.0015364 Fitz, D., Jakschitz, T. & Rode, B. M. in Origins of Life: The Primal Self-Organization (eds. Egel, R., Lankenau, D.-H. & Mulkidjanian, A. Y.), pp. 109–127 (Springer Berlin Heidelberg, 2011). Forterre, P. & Prangishvili, D. The origin of viruses // Research in Microbiology, 2009, vol. 160, pp. 466–472. DOI: 16/j.resmic.2009.07.008 Forterre, P. A new fusion hypothesis for the origin of Eukarya: better than previous ones, but probably also wrong // Research in microbiology, 2011, vol. 162, pp. 77–91. DOI: 10.1016/j.resmic.2010.10.005 Forterre, P. The origin of DNA genomes and DNA replication proteins // Current Opinion in Microbiology, 2002, vol. 5, pp. 525–532. DOI: 10.1016/S1369–5274(02)00360–0 Forterre, P. The two ages of the RNA world, and the transition to the DNA world: a story of viruses and cells // Biochimie, 2005, vol. 87, pp. 793–803. DOI: 10.1016/j. biochi.2005.03.015 Forterre, P. Three RNA cells for ribosomal lineages and three DNA viruses to replicate their genomes: A hypothesis for the origin of cellular domain // Proceedings of the National Academy of Sciences of the United States of America, 2006, vol. 103, pp. 3669–3674. DOI: 10.1073/pnas.0510333103 Fuerst, J. A. & Sagulenko, E. Keys to eukaryality: planctomycetes and ancestral evolution of cellular complexity // Frontiers in microbiology, 2012, vol. 3, pp. 167. DOI: 10.3389/fmicb.2012.00167 Giulio, M. D. A Comparison Among the Models Proposed to Explain the Origin of the tRNA Molecule: A Synthesis // Journal of Molecular Evolution, 2009, vol. 69, pp. 1–9. DOI: 10.1007/s00239-009-9248-z Gomes, R., Levison, H. F., Tsiganis, K. & Morbidelli, A. Origin of the cataclysmic Late Heavy Bombardment period of the terrestrial planets // Nature, 2005, vol. 435, pp. 466–469. DOI: 10.1038/nature03676 Grein, F., Ramos, A. R., Venceslau, S. S. & Pereira, I. A. C. Unifying concepts in anaerobic respiration: Insights from dissimilatory sulfur metabolism // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2013, vol. 1827, pp. 145–160. DOI: 10.1016/j.bbabio.2012.09.001 Gribaldo, S., Talla, E. & Brochier-Armanet, C. Evolution of the haem copper oxidases superfamily: a rooting tale // Trends in Biochemical Sciences, 2009, vol. 34, pp. 375–381. DOI: 10.1016/j.tibs.2009.04.002 Gupta, R. S. Origin and Spread of Photosynthesis Based upon Conserved Sequence Features in Key Bacteriochlorophyll Biosynthesis Proteins // Molecular Biology and Evolution, 2012, vol. 29, pp. 3397–3412. DOI: 10.1093/molbev/mss145 Guzman, M. I. & Martin, S. T. Prebiotic metabolism: production by mineral photoelectrochemistry of alpha-ketocarboxylic acids in the reductive tricarboxylic acid cycle // Astrobiology, 2009, vol. 9, pp. 833–842. DOI: 10.1089/ast.2009.0356 Hagan, W. J. Uracil-Catalyzed Synthesis of Acetyl Phosphate: A Photochemical Driver for Protometabolism // ChemBioChem, 2010, vol. 11, pp. 383–387. DOI: 10.1002/cbic. 200900433 Haines, T. H. Do sterols reduce proton and sodium leaks through lipid bilayers? // Progress in lipid research, 2001, vol. 40, pp. 299–324. DOI: 10.1016/S0163–7827(01)00009–1 Halliday, A. N. The Origin and Earliest History of the Earth // Planets, Asteriods, Comets and The Solar System, 2014, vol. 1, pp. 149–211. Halliday, A. N. The origins of volatiles in the terrestrial planets // Geochimica et Cosmochimica Acta, 2013, vol. 105, pp. 146–171. DOI: 10.1016/j.gca.2012.11.015 Harish, A. & Caetano-Anollé s, G. Ribosomal History Reveals Origins of Modern Protein Synthesis // PLoS ONE, 2012, vol. 7, p. e32776. DOI: 10.1371/journal.pone.0032776 Hartman, H. & Smith, T. F. The Evolution of the Ribosome and the Genetic Code // Life, 2014, vol. 4, pp. 227–249. DOI: 10.3390/life4020227 Hazen, R. M., Filley, T. R. & Goodfriend, G. A. Selective adsorption of l– and d-amino acids on calcite: Implications for biochemical homochirality // Proceedings of the National Academy of Sciences, 2001, vol. 98, pp. 5487–5490. DOI: 10.1073/pnas.101085998 Hein, J. E. & Blackmond, D. G. On the Origin of Single Chirality of Amino Acids and Sugars in Biogenesis // Accounts of chemical research, 2012. DOI: 10.1021/ar200316n Hohmann-Marriott, M. F. & Blankenship, R. E. Evolution of photosynthesis // Annual Review of Plant Biology, 2011, vol. 62, pp. 515–548. DOI: 10.1146/annurev-arplant-042110–103811 Hohn, T., Hohn, B. & Pfeiffer, P. Reverse transcription in CaMV // Trends in Biochemical Sciences, 1985, vol. 10, pp. 205–209. DOI: 10.1016/0968–0004(85)90193–8 Hsiao, C. et al. Molecular paleontology: a biochemical model of the ancestral ribosome // Nucleic Acids Research, 2013 gkt023. DOI: 10.1093/nar/gkt023 Hsiao, C., Mohan, S., Kalahar, B. K. & Williams, L. D. Peeling the Onion: Ribosomes Are Ancient Molecular Fossils // Molecular Biology and Evolution, 2009, vol. 26, pp. 2415–2425. DOI: 10.1093/molbev/msp163 Huang, W. & Ferris, J. P. One-step, regioselective synthesis of up to 50-mers of RNA oligomers by montmorillonite catalysis // Journal of the American Chemical Society, 2006, vol. 128, pp. 8914–8919. DOI: 10.1021/ja061782k Ii, H. J. C. & Bada, J. L. in Genesis – In The Beginning (ed. Seckbach, J.), pp. 3–33 (Springer Netherlands, 2012). Inagaki, F. et al. Microbial community in a sediment-hosted CO2 lake of the southern Okinawa Trough hydrothermal system // Proceedings of the National Academy of Sciences, 2006, vol. 103, pp. 14164–14169. DOI: 10.1073/pnas.0606083103 Ingar, A.-A., Luke, R. W. A., Hayter, B. R. & Sutherland, J. D. Synthesis of Cytidine Ribonucleotides by Stepwise Assembly of the Heterocycle on a Sugar Phosphate // ChemBioChem, 2003, vol. 4, pp. 504–507. DOI: 10.1002/cbic. 200300554 Iyer, L. M., Koonin, E. V. & Aravind, L. Evolutionary connection between the catalytic subunits of DNA-dependent RNA polymerases and eukaryotic RNA-dependent RNA polymerases and the origin of RNA polymerases // BMC Structural Biology, 2003, vol. 3, p. 1. DOI: 10.1186/1472-6807-3-1 Jadhav, V. R. & Yarus, M. Acyl-CoAs from Coenzyme Ribozymes // Biochemistry, 2002, vol. 41, pp. 723–729. DOI: 10.1021/bi011803h Jadhav, V. R. & Yarus, M. Coenzymes as coribozymes // Biochimie, 2002, vol. 84, pp. 877–888. DOI: 10.1016/S0300–9084(02)01404–9 Jheeta, S. & Joshi, P. C. Prebiotic RNA Synthesis by Montmorillonite Catalysis // Life: Open Access Journal, 2014, vol. 4, pp. 318–330. DOI: 10.3390/life4030318 Kaula, W. M. Constraints on Venus Evolution from Radiogenic Argon // Icarus, 1999, vol. 139, pp. 32–39. DOI: 10.1006/icar. 1999.6082 Keeling, P. J. Diversity and evolutionary history of plastids and their hosts // American Journal of Botany, 2004, vol. 91, pp. 1481–1493. DOI: 10.3732/ajb.91.10.1481 Keller, M. A., Turchyn, A. V. & Ralser, M. Non-enzymatic glycolysis and pentose phosphate pathway-like reactions in a plausible Archean ocean // Molecular Systems Biology, 2014, vol. 10. DOI: 10.1002/msb.20145228 Kim, H.-J. et al. Synthesis of Carbohydrates in Mineral-Guided Prebiotic Cycles // Journal of the American Chemical Society, 2011, vol. 133, pp. 9457–9468. DOI: 10.1021/ja201769f Koch, A. L. & Silver, S. in Advances in Microbial Physiology (ed. Robert K. Poole), vol. 50, pp. 227–259 (Academic Press, 2005). Kofoed, J., Reymond, J.-L. & Darbre, T. Prebiotic carbohydrate synthesis: zinc-proline catalyzes direct aqueous aldol reactions of alpha-hydroxy aldehydes and ketones // Organic & biomolecular chemistry, 2005, vol. 3, pp. 1850–1855. DOI: 10.1039/b501512j Koonin, E. V. & Novozhilov, A. S. Origin and evolution of the genetic code: the universal enigma // IUBMB Life, 2009, vol. 61, pp. 99–111. DOI: 10.1002/iub.146 Koonin, E. V. On the Origin of Cells and Viruses // Annals of the New York Academy of Sciences, 2009, vol. 1178, pp. 47–64. DOI: 10.1111/j.1749–6632.2009.04992.x Koonin, E. V. Temporal order of evolution of DNA replication systems inferred by comparison of cellular and viral DNA polymerases // Biology Direct, 2006, Vol. 1, pp. 39. DOI: 10.1186/1745-6150-1-39 Kreysing, M., Keil, L., Lanzmich, S. & Braun, D. Heat flux across an open pore enables the continuous replication and selection of oligonucleotides towards increasing length // Nature Chemistry, 2015, vol. 7, pp. 203–208. DOI: 10.1038/nchem.2155 Kritsky, M. S., Kolesnikov, M. P. & Telegina, T. A. Modeling of abiogenic synthesis of ATP // Doklady Biochemistry and Biophysics, 2007, vol. 417, pp. 313–315. DOI: 10.1134/S1607672907060063 Lamb, D. C. et al. Lanosterol Biosynthesis in the Prokaryote Methylococcus Capsulatus: Insight into the Evolution of Sterol Biosynthesis // Molecular Biology and Evolution, 2007, vol. 24, pp. 1714–1721. DOI: 10.1093/molbev/msm090 Laskar, J. & Robutel, P. The chaotic obliquity of the planets // Nature, 1993, vol. 361, pp. 608–612. DOI: 10.1038/361608a0 Leipe, D. D., Aravind, L. & Koonin, E. V. Did DNA replication evolve twice independently? // Nucleic Acids Research, 1999, vol. 27, pp. 3389–3401. DOI: 10.1093/nar/27.17.3389. Lincoln, T. A. & Joyce, G. F. Self-sustained Replication of an RNA Enzyme // Science (New York, N. Y.), 2009, vol. 323, pp. 1229–1232. DOI: 10.1126/science.1167856 Lissauer, J. J. et al. A closely packed system of low-mass, low-density planets transiting Kepler-11 // Nature, 2011, vol. 470, pp. 53–58 (2011). DOI: 10.1038/nature09760 Liu, R. & Orgel, L. E. Enzymatic synthesis of polymers containing nicotinamide mononucleotide // Nucleic Acids Research, 1995, vol. 23, pp, 3742–3749. DOI: 10.1093/nar/23.18.3742 Lombard, J., Ló pez-Garcí a, P. & Moreira, D. The early evolution of lipid membranes and the three domains of life // Nature Reviews Microbiology, 2012, vol. 10, pp. 507–515. DOI: 10.1038/nrmicro2815 Macnaughton, T. B., Shi, S. T., Modahl, L. E. & Lai, M. M. C. Rolling Circle Replication of Hepatitis Delta Virus RNA Is Carried Out by Two Different Cellular RNA Polymerases // Journal of Virology, 2002, vol. 76, pp. 3920–3927. DOI: 10.1128/JVI.76.8.3920–3927.2002 Makarova, K. S., Yutin, N., Bell, S. D. & Koonin, E. V. Evolution of diverse cell division and vesicle formation systems in Archaea // Nature reviews. Microbiology, 2010, vol. 8, pp. 731–741. DOI: 10.1038/nrmicro2406 Malyshev, D. A. et al. A semi-synthetic organism with an expanded genetic alphabet // Nature, 2014, advance online publication. DOI: 10.1038/nature13314 Margulis, L., Chapman, M., Guerrero, R. & Hall, J. The last eukaryotic common ancestor (LECA): Acquisition of cytoskeletal motility from aerotolerant spirochetes in the Proterozoic Eon // Proceedings of the National Academy of Sciences, 2006, vol. 103, pp. 13080–13085. DOI: 10.1073/pnas.0604985103 Marreiros, B. C., Batista, A. P., Duarte, A. M. S. & Pereira, M. M. A missing link between complex I and group 4 membrane-bound [NiFe] hydrogenases // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2013, vol. 1827, pp. 198–209. DOI: 10.1016/j.bbabio.2012.09.012 Martin, H. et al. 4. Building of a Habitable Planet // Earth, Moon, and Planets, 2006, vol. 98, pp. 97–151. DOI: 10.1007/s11038-006-9088-4 Martin, L. L., Unrau, P. J. & Mü ller, U. F. RNA Synthesis by in Vitro Selected Ribozymes for Recreating an RNA World // Life, 2015, vol. 5, pp. 247–268. DOI: 10.3390/life5010247 Martin, W. & Russell, M. J. On the origin of biochemistry at an alkaline hydrothermal vent // Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 2007, vol. 362, pp. 1887–1925. DOI: 10.1098/rstb.2006.1881 Marty, B. The origins and concentrations of water, carbon, nitrogen and noble gases on Earth // Earth and Planetary Science Letters, 2012, vol. 313–314, pp. 56–66. DOI: 10.1016/j.epsl.2011.10.040 Mast, C. B., Schink, S., Gerland, U. & Braun, D. Escalation of polymerization in a thermal gradient // Proceedings of the National Academy of Sciences, 2013, vol. 110, pp. 8030–8035. DOI: 10.1073/pnas.1303222110 Mellersh, A. R. A model for the prebiotic synthesis of peptides which throws light on the origin of the genetic code and the observed chirality of life // Origins of life and evolution of the biosphere, 1993, vol. 23, pp. 261–274. DOI: 10.1007/BF01581903 Milbury, C., Schubert, G., Raymond, C. A., Smrekar, S. E. & Langlais, B. The history of Mars' dynamo as revealed by modeling magnetic anomalies near Tyrrhenus Mons and Syrtis Major // Journal of Geophysical Research: Planets, 2012, vol. 117, E10007. DOI: 10.1029/2012JE004099 Mittapalli, G. K. et al. Mapping the Landscape of Potentially Primordial Informational Oligomers: Oligodipeptides Tagged with 2, 4-Disubstituted 5-Aminopyrimidines as Recognition Elements // Angewandte Chemie International Edition, 2007, vol. 46, pp. 2478–2484. DOI: 10.1002/anie.200603209 Mix, L., Haig, D. & Cavanaugh, C. Phylogenetic Analyses of the Core Antenna Domain: Investigatingthe Origin of Photosystem I // Journal of Molecular Evolution, 2005, vol. 60, pp. 153–163. DOI: 10.1007/s00239-003-0181-2 Mö ller, W. & Janssen, G. M. C. Transfer RNAs for primordial amino acids contain remnants of a primitive code at position 3 to 5 // Biochimie, 1990, vol. 72, pp. 361–368. DOI: 10.1016/0300–9084(90)90033-D Montmerle, T. et al. 3. Solar System Formation and Early Evolution: the First 100 Million Years // Earth, Moon, and Planets, 2006, vol. 98, pp. 39–95. DOI: 10.1007/s11038-006-9087-5 Moore, S. J. et al. Elucidation of the anaerobic pathway for the corrin component of cobalamin (vitamin B12) // Proceedings of the National Academy of Sciences, 2013, vol. 110, pp. 14906–14911. DOI: 10.1073/pnas.1308098110 Morbidelli, A., Brasser, R., Gomes, R., Levison, H. F. & Tsiganis, K. Evidence from the Asteroid Belt for a Violent Past Evolution of Jupiter's Orbit // The Astronomical Journal, 2010, vol. 140, p. 1391. DOI: 10.1088/0004–6256/140/5/1391 Morbidelli, A., Levison, H. F., Tsiganis, K. & Gomes, R. Chaotic capture of Jupiter's Trojan asteroids in the early Solar System // Nature, 2005, vol. 435, pp. 462–465. DOI: 10.1038/nature03540 Moreira, M. Noble Gas Constraints on the Origin and Evolution of Earth's Volatiles // Geochemical Perspectives, 2013, vol. 2, pp. 229–230. URL: http: //perspectives.geoscienceworld.org/content/2/2/229 Mulkidjanian, A. Y. & Junge, W. On the origin of photosynthesis as inferred from sequence analysis // Photosynthesis Research, 1997, vol. 51, pp. 27–42. DOI: 10.1023/A: 1005726809084 Mulkidjanian, A. Y. & Galperin, M. Y. On the origin of life in the zinc world. 2. Validation of the hypothesis on the photosynthesizing zinc sulfide edifices as cradles of life on Earth // Biology Direct, 2009, vol. 4, p. 27. DOI: 10.1186/1745-6150-4-27 Mulkidjanian, A. Y. & Galperin, M. Y. Physico-chemical and evolutionary constraints for the formation and selection of first biopolymers: towards the consensus paradigm of the abiogenic origin of life // Chemistry & Biodiversity, 2007, vol. 4, pp. 2003–2015. DOI: 10.1002/cbdv.200790167 Mulkidjanian, A. Y. et al. The cyanobacterial genome core and the origin of photosynthesis // Proceedings of the National Academy of Sciences, 2006, vol. 103, pp. 13126–13131. DOI: 10.1073/pnas. 0605709103 Mulkidjanian, A. Y. On the origin of life in the zinc world: 1. Photosynthesizing, porous edifices built of hydrothermally precipitated zinc sulfide as cradles of life on Earth // Biology Direct, 2009, vol. 4, p. 26. DOI: 10.1186/1745-6150-4-26 Mulkidjanian, A. Y., Bychkov, A. Y., Dibrova, D. V., Galperin, M. Y. & Koonin, E. V. Origin of first cells at terrestrial, anoxic geothermal fields // Proceedings of the National Academy of Sciences, 2012, vol. 109, pp. E821 – E830. DOI: 10.1073/pnas.1117774109 Mulkidjanian, A. Y., Cherepanov, D. A. & Galperin, M. Y. Survival of the fittest before the beginning of life: selection of the first oligonucleotide-like polymers by UV light // BMC Evolutionary Biology, 2003, vol. 3, p. 12. DOI: 10.1186/1471-2148-3-12 Mulkidjanian, A. Y., Galperin, M. Y. & Koonin, E. V. Co-evolution of primordial membranes and membrane proteins // Trends in Biochemical Sciences, 2009, vol. 34, pp. 206–215. DOI: 10.1016/j.tibs.2009.01.005 Mulkidjanian, A. Y., Galperin, M. Y., Makarova, K. S., Wolf, Y. I. & Koonin, E. V. Evolutionary primacy of sodium bioenergetics // Biology Direct, 2008, vol. 3, p. 13. DOI: 10.1186/1745-6150-3-13 Navarro-Gonzá lez, R., McKay, C. P. & Mvondo, D. N. A possible nitrogen crisis for Archaean life due to reduced nitrogen fixation by lightning // Nature, 2001, Proceedings of the National Academy of Sciences, 2012, vol. 109, pp. 61–64. DOI: 10.1038/35083537 Nelson, K. E., Levy, M. & Miller, S. L. Peptide nucleic acids rather than RNA may have been the first genetic molecule // Proceedings of the National Academy of Sciences of the United States of America 2000, vol. 97, pp. 3868–3871. DOI: 10.1073/pnas.97.8.3868 Nemchin, A. A. et al. A light carbon reservoir recorded in zircon-hosted diamond from the Jack Hills // Nature, 2008, vol. 454, pp. 92–95. DOI: 10.1038/nature07102 Nielsen, P. E. Peptide Nucleic Acids and the Origin of Life // Chemistry & Biodiversity, 2007, vol. 4, pp. 1996–2002. DOI: 10.1002/cbdv.200790166 Noller, H. F. Evolution of Protein Synthesis from an RNA World // Cold Spring Harbor Perspectives in Biology, 2012, vol. 4, p. a003681. DOI: 10.1101/cshperspect.a003681 Noller, H. F. The driving force for molecular evolution of translation // RNA, 2004, vol. 10, pp. 1833–1837. DOI: 10.1261/rna.7142404 Nunoura, T. et al. Insights into the evolution of Archaea and eukaryotic protein modifier systems revealed by the genome of a novel archaeal group // Nucleic Acids Research, 2011, vol. 39, pp. 3204–3223. DOI: 10.1093/nar/gkq1228 Ogasawara, H. et al. Synthesizing Oligomers from Monomeric Nucleotides in Simulated Hydrothermal Environments // Origins of life and evolution of the biosphere, 2000, vol. 30, pp. 519–526. DOI: 10.1023/A: 1026539708173 Olson, J. & Blankenship, R. Thinking About the Evolution of Photosynthesis // Photosynthesis Research, 2004, vol. 80, pp. 373–386. DOI: 10.1023/B: PRES.0000030457.06495.83 Orgel, L. E. The Implausibility of Metabolic Cycles on the Prebiotic Earth // PLoS Biology, 2008, vol. 6. DOI: 10.1371/journal.pbio.0060018 Otroshchenko, V. A. & Vasilyeva, N. V. Formation of RNA oligonucleotides over the mineral surface preliminary irradiated with UV light // Reaction Kinetics and Catalysis Letters, 2009, vol. 97, pp. 151–156. DOI: 10.1007/s11144-009-0006-4 Outten, F. W. Iron-sulfur clusters as oxygen-responsive molecular switches // Nature Chemical Biology, 2007, vol. 3, pp. 206–207. DOI: 10.1038/nchembio0407–206 Padmanabhan, S., York, E. J., Stewart, J. M. & Baldwin, R. L. Helix propensities of basic amino acids increase with the length of the side-chain // Journal of Molecular Biology, 1996, vol. 257, pp. 726–734. DOI: 10.1006/jmbi.1996.0197 Pasek, M. A. Rethinking early Earth phosphorus geochemistry // Proceedings of the National Academy of Sciences, 2008, vol. 105, pp. 853–858. DOI: 10.1073/pnas.0708205105 Patel, S. C., Bradley, L. H., Jinadasa, S. P. & Hecht, M. H. Cofactor binding and enzymatic activity in an unevolved superfamily of de novo designed 4-helix bundle proteins // Protein Science, 2009, vol. 18, pp. 1388–1400. DOI: 10.1002/pro.147 Pearson, A., Budin, M. & Brocks, J. J. Phylogenetic and biochemical evidence for sterol synthesis in the bacterium Gemmata obscuriglobus // Proceedings of the National Academy of Sciences, 2003, vol. 100, pp. 15352–15357. DOI: 10.1073/pnas.2536559100 Pech, H. et al. Detection of Geothermal Phosphite Using High-Performance Liquid Chromatography // Environmental Science & Technology, 2009, vol. 43, pp. 7671–7675. DOI: 10.1021/es901469t Pei, J., Li, W., Kinch, L. N. & Grishin, N. V. Conserved evolutionary units in the heme-copper oxidase superfamily revealed by novel homologous protein families // Protein Science, 2014, vol. 23, pp. 1220–1234. DOI: 10.1002/pro.2503 Pestunova, O., Simonov, A., Snytnikov, V., Stoyanovsky, V. & Parmon, V. Putative mechanism of the sugar formation on prebiotic Earth initiated by UV-radiation // Advances in Space Research, 2005, vol. 36, pp. 214–219. DOI: 10.1016/j.asr.2005.02.049 Philippot, P. et al. Early Archaean Microorganisms Preferred Elemental Sulfur, Not Sulfate // Science, 2007, vol. 317, pp. 1534–1537. DOI: 10.1126/science.1145861 Philippot, P., van Zuilen, M. & Rollion-Bard, C. Variations in atmospheric sulphur chemistry on early Earth linked to volcanic activity // Nature Geoscience, 2012, vol. 5, pp. 668–674. DOI: 10.1038/ngeo1534 Poole, A., Penny, D. & Sjö berg, B.-M. Methyl-RNA: an evolutionary bridge between RNA and DNA? // Chemistry & Biology, 2000, vol. 7, pp. R207 – R216. DOI: 10.1016/S1074–5521(00)00042–9 Poole, A. M. & Logan, D. T. Modern mRNA Proofreading and Repair: Clues that the Last Universal Common Ancestor Possessed an RNA Genome? // Molecular Biology and Evolution, 2005, vol. 22, pp. 1444–1455. DOI: 10.1093/molbev/msi132 Powner, M. W., Gerland, B. & Sutherland, J. D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions // Nature, 2009, vol. 459, pp. 239–242. DOI: 10.1038/nature08013 Pugachev, K. V. et al. High Fidelity of Yellow Fever Virus RNA Polymerase // Journal of Virology, 2004, vol. 78, pp. 1032–1038. DOI: 10.1128/JVI.78.2.1032–1038.2004 Quayle, J. R. & Ferenci, T. Evolutionary aspects of autotrophy // Microbiological Reviews, 1978, vol. 42, pp. 251–273. Rajamani, S. et al. Lipid-assisted Synthesis of RNA-like Polymers from Mononucleotides // Origins of Life and Evolution of Biospheres, 2007, vol. 38, pp. 57–74. DOI: 10.1007/s11084-007-9113-2 Ricardo, A., Carrigan, M. A., Olcott, A. N. & Benner, S. A. Borate Minerals Stabilize Ribose // Science, 2004, vol. 303, pp. 196–196. DOI: 10.1126/science.1092464 Riguzzi, F., Panza, G., Varga, P. & Doglioni, C. Can Earth's rotation and tidal despinning drive plate tectonics? // Tectonophysics, 2010, vol. 484, pp. 60–73. DOI: 10.1016/j.tecto.2009.06.012 Ritson, D. & Sutherland, J. D. Prebiotic synthesis of simple sugars by photoredox systems chemistry // Nature chemistry, 2012, vol. 4, pp. 895–899. DOI: 10.1038/nchem.1467 Robertson, M. P., Knudsen, S. M. & Ellington, A. D. In vitro selection of ribozymes dependent on peptides for activity // RNA, 2004, vol. 10, pp. 114–127. DOI: 10.1261/rna.5900204 Rodin, A. S., Szathmá ry, E. & Rodin, S. N. On origin of genetic code and tRNA before translation // Biology Direct, 2011, vol. 6, p. 14. DOI: 10.1186/1745-6150-6-14 Rodin, A. S., Szathmá ry, E. & Rodin, S. N. One ancestor for two codes viewed from the perspective of two complementary modes of tRNA aminoacylation // Biology Direct, 2009, vol. 4, p. 4. DOI: 10.1186/1745-6150-4-4 Rodin, S. N. & Ohno, S. Two types of aminoacyl-trna synthetases could be originally encoded by complementary strands of the same nucleic ACID // Origins of life and evolution of the biosphere, 1995, vol. 25, pp. 565–589. DOI: 10.1007/BF01582025 Root-Bernstein, M. & Root-Bernstein, R. The ribosome as a missing link in the evolution of life // Journal of Theoretical Biology, 2015, vol. 367, pp. 130–158. DOI: 10.1016/j.jtbi.2014.11.025 Roth, A. & Breaker, R. R. An amino acid as a cofactor for a catalytic polynucleotide // Proceedings of the National Academy of Sciences, 1998, vol. 95, pp. 6027–6031. Rothschild, L. J. The evolution of photosynthesis… again? // Philosophical Transactions of the Royal Society of London B: Biological Sciences, 2008, vol. 363, pp. 2787–2801. DOI: 10.1098/rstb.2008.0056 Russell, M. J. et al. The Drive to Life on Wet and Icy Worlds // Astrobiology, 2014, vol. 14, pp. 308–343. DOI: 10.1089/ast.2013.1110 Ryu, Y., Kim, K.-J., Roessner, C. A. & Scott, A. I. Decarboxylative Claisen condensation catalyzed by in vitro selected ribozymes // Chemical Communications, 2006, vol. 13, pp. 1439–1441. DOI: 10.1039/b517160a Saladino, R., Crestini, C., Ciciriello, F., Costanzo, G. & Di Mauro, E. Formamide Chemistry and the Origin of Informational Polymers // Chemistry & Biodiversity, 2007, vol. 4, pp. 694–720. DOI: 10.1002/cbdv.200790059 Sato, T., Atomi, H. & Imanaka, T. Archaeal Type III RuBisCOs Function in a Pathway for AMP Metabolism // Science, 2007, vol. 315, pp. 1003–1006. DOI: 10.1126/science.1135999 Schoepp-Cothenet, B. et al. On the universal core of bioenergetics // Biochimica et Biophysica Acta (BBA) – Bioenergetics, 2013, vol. 1827, pp. 79–93. DOI: 10.1016/j. bbabio.2012.09.005 Schö ning, K.-U. et al. Chemical Etiology of Nucleic Acid Structure: The α -Threofuranosyl– (3'→ 2') Oligonucleotide System // Science, 2000, vol. 290, pp. 1347–1351. DOI: 10.1126/science.290.5495.1347 Sczepanski, J. T. & Joyce, G. F. A cross-chiral RNA polymerase ribozyme // Nature, 2014, vol. 515, pp. 440–442. DOI: 10.1038/nature13900 Sen, D. & Poon, L. C. H. RNA and DNA complexes with hemin [Fe (III) heme] are efficient peroxidases and peroxygenases: how do they do it and what does it mean? // Critical Reviews in Biochemistry and Molecular Biology, 2011, vol. 46, pp. 478–492. DOI: 10.3109/10409238.2011.618220 Senanayake, S. D. & Idriss, H. Photocatalysis and the origin of life: synthesis of nucleoside bases from formamide on TiO2 (001) single surfaces // Proceedings of the National Academy of Sciences of the United States of America, 2006, vol. 103, pp. 1194–1198. DOI: 10.1073/pnas.0505768103 Shimada, H. & Yamagishi, A. Stability of Heterochiral Hybrid Membrane Made of Bacterial sn-G3P Lipids and Archaeal sn-G1P Lipids // Biochemistry, 2011, vol. 50, pp. 4114–4120. DOI: 10.1021/bi200172d Shimizu, M., Yamagishi, A., Kinoshita, K., Shida, Y. & Oshima, T. Prebiotic Origin of Glycolytic Metabolism: Histidine and Cysteine can Produce Acetyl CoA from Glucose via Reactions Homologous to Non-phosphorylated Entner-Doudoroff Pathway // Journal of Biochemistry, 2008, vol. 144, pp. 383–388. DOI: 10.1093/jb/mvn073 Silverman, S. K. Deoxyribozymes: DNA catalysts for bioorganic chemistry // Organic & Biomolecular Chemistry, 2004, vol. 2, pp. 2701–2706. DOI: 10.1039/B411910J Sivan, O., Antler, G., Turchyn, A. V., Marlow, J. J. & Orphan, V. J. Iron oxides stimulate sulfate-driven anaerobic methane oxidation in seeps // Proceedings of the National Academy of Sciences, 2014, vol. 111, pp. E4139 – E4147. DOI: 10.1073/pnas.1412269111 Smith, E. & Morowitz, H. J. Universality in intermediary metabolism // Proceedings of the National Academy of Sciences of the United States of America, 2004, vol. 101, pp. 13168–13173. DOI: 10.1073/pnas.0404922101 Solov'yov, I. A. & Schulten, K. Magnetoreception through Cryptochrome May Involve Superoxide // Biophysical Journal, 2009, vol. 96, pp. 4804–4813. DOI: 10.1016/j.bpj.2009.03.048 Sousa, F. L. et al. Early bioenergetic evolution // Philosophical Transactions of the Royal Society B: Biological Sciences, 2013, vol. 368, Article ARTN 20130088. DOI: 10.1098/rstb.2013.0088 Spang, A. et al. Complex archaea that bridge the gap between prokaryotes and eukaryotes // Nature advance online publication (2015). DOI: 10.1038/nature14447 Szabó -Nagy, A. & Keszthelyi, L. Demonstration of the parity-violating energy difference between enantiomers // Proceedings of the National Academy of Sciences, 1999, vol. 96, pp. 4252–4255. DOI: 10.1073/pnas.96.8.4252 Szathmá ry, E. Coding coenzyme handles: a hypothesis for the origin of the genetic code // Proceedings of the National Academy of Sciences, 1993, vol. 90, pp. 9916–9920. DOI: 10.1073/pnas.90.21.9916 Szostak, J. W. The eightfold path to non-enzymatic RNA replication // Journal of Systems Chemistry, 2012, vol. 3, p. 2. DOI: 10.1186/1759-2208-3-2 Takemura, M. Poxviruses and the origin of the eukaryotic nucleus // Journal of molecular evolution, 2001, vol. 52, pp. 419–425. DOI: 10.1007/s002390010171 Takemura, M., Yokobori, S. & Ogata, H. Evolution of Eukaryotic DNA Polymerases via Interaction Between Cells and Large DNA Viruses // Journal of Molecular Evolution, 2015, vol. 81, pp. 24–33. DOI: 10.1007/s00239-015-9690-z Tang, K.-H. et al. Complete genome sequence of the filamentous anoxygenic phototrophic bacterium Chloroflexus aurantiacus // BMC Genomics, 2011, vol. 12, p. 334. DOI: 10.1186/1471-2164-12-334 Tang, K.-H., Yue, H. & Blankenship, R. E. Energy metabolism of Heliobacterium modesticaldum during phototrophic and chemotrophic growth // BMC Microbiology, 2010, vol. 10, p. 150. DOI: 10.1186/1471-2180-10-150 Telegina, T. A., Kolesnikov, M. P., Vechtomova, Y. L., Buglak, A. A. & Kritsky, M. S. Abiotic photophosphorylation model based on abiogenic flavin and pteridine pigments // Journal of molecular evolution, 2013, vol. 76, pp. 332–342. DOI: 10.1007/s00239-013-9562-3 The Limits of Organic Life in Planetary Systems. Available at: http: //books.nap.edu/openbook.php? record_id=11919& page=69. (Accessed: 3rd September 2015). Tielens, A. G. M., Rotte, C., van Hellemond, J. J. & Martin, W. Mitochondria as we don't know them // Trends in Biochemical Sciences, 2002, vol. 27, pp. 564–572. DOI: 16/S0968–0004(02)02193-X Tsiganis, K., Gomes, R., Morbidelli, A. & Levison, H. F. Origin of the orbital architecture of the giant planets of the Solar System // Nature, 2005, vol. 435, pp. 459–461. DOI: 10.1038/nature03539 Tsukiji, S., Pattnaik, S. B. & Suga, H. Reduction of an Aldehyde by a NADH/Zn2+-Dependent Redox Active Ribozyme // Journal of the American Chemical Society, 2004, vol. 126, pp. 5044–5045. DOI: 10.1021/ja0495213 Turick, C. E., Ekechukwu, A. A., Milliken, C. E., Casadevall, A. & Dadachova, E. Gamma radiation interacts with melanin to alter its oxidation – reduction potential and results in electric current production // Bioelectrochemistry, 2011, vol. 82, pp. 69–73. DOI: 10.1016/j.bioelechem.2011.04.009 Vakulenko, S. & Grigoriev, D. Evolution in random environment and structural instability // Journal of Mathematical Sciences, 2006, vol. 138, pp. 5644–5662. DOI: 10.1007/s10958-006-0333-1 Van Flandern, T. C. & Harrington, R. S. A dynamical investigation of the conjecture that Mercury is an escaped satellite of Venus // Icarus, 1976, vol. 28, pp. 435–440. DOI: 10.1016/0019–1035(76)90116–0 Viedma, C., Ortiz, J. E., Torres, T. de, Izumi, T. & Blackmond, D. G. Evolution of Solid Phase Homochirality for a Proteinogenic Amino Acid // Journal of the American Chemical Society, 2008, vol. 30, pp. 15274–15275. DOI: 10.1021/ja8074506 Vlassov, A. V., Johnston, B. H., Landweber, L. F. & Kazakov, S. Ligation activity of fragmented ribozymes in frozen solution: implications for the RNA world // Nucleic Acids Research, 2004, vol. 2, pp. 2966–2974. DOI: 10.1093/nar/gkh601
Wacey, D., McLoughlin, N., Whitehouse, M. J. & Kilburn, M. R. Two coexisting sulfur metabolisms in a ca. 3400 Ma sandstone // Geology, 2010, vol. 38, pp. 1115–1118. DOI: 10.1130/G31329.1 Wä chtershä user, G. From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya // Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 2006, vol. 361, pp. 1787–1806; discussion 1806–1808. DOI: 10.1098/rstb.2006.1904 Weber, A. L. & Miller, S. L. Reasons for the occurrence of the twenty coded protein amino acids // Journal of Molecular Evolution, 1981, vol. 17, pp. 273–284. DOI: 10.1007/BF01795749 Williams, G. E. Geological constraints on the Precambrian history of Earth's rotation and the Moon's orbit // Reviews of Geophysics, 2000, vol. 38, pp. 37–59. DOI: 10.1029/1999RG900016 Wochner, A., Attwater, J., Coulson, A. & Holliger, P. Ribozyme-Catalyzed Transcription of an Active Ribozyme // Science, 2011, vol. 32, pp. 209–212. DOI: 10.1126/science.1200752 Wray, J. J. et al. Prolonged magmatic activity on Mars inferred from the detection of felsic rocks // Nature Geoscience, 2013, vol. 6, pp. 1013–1017. DOI: 10.1038/ngeo1994 Yu, H., Zhang, S. & Chaput, J. C. Darwinian evolution of an alternative genetic system provides support for TNA as an RNA progenitor // Nature Chemistry, 2012, vol. 4, pp. 183–187. DOI: 10.1038/nchem.1241 Yutin, N. & Koonin, E. V. Archaeal origin of tubulin // Biology direct, 2012, vol. 7, p. 10. DOI: 10.1186/1745-6150-7-10 Yutin, N., Wolf, M. Y., Wolf, Y. I. & Koonin, E. V. The origins of phagocytosis and eukaryogenesis // Biology direct, 2009, vol. 4, p. 9. DOI: 10.1186/1745-6150-4-9 Zahnle, K. et al. Emergence of a Habitable Planet // Space Science Reviews, 2007, vol. 129, pp. 35–78. DOI: 10.1007/s11214-007-9225-z Zahnle, K., Schaefer, L. & Fegley, B. Earth's Earliest Atmospheres // Cold Spring Harbor Perspectives in Biology, 2010, vol. 2. DOI: 10.1101/cshperspect.a004895 Zarzycki, J., Brecht, V., Mü ller, M. & Fuchs, G. Identifying the missing steps of the autotrophic 3-hydroxypropionate CO2 fixation cycle in Chloroflexus aurantiacus // Proceedings of the National Academy of Sciences, 2009, vol. 106, pp. 21317–21322. DOI: 10.1073/pnas.0908356106 Zhang, L., Peritz, A. & Meggers, E. A Simple Glycol Nucleic Acid // Journal of the American Chemical Society, 2005, vol. 127, pp. 4174–4175. DOI: 10.1021/ja042564z
Сноски 1 Наглядное представление об устойчивости планетных орбит можно получить в онлайн-игре Super Planet Crash (http: //www.stefanom.org/spc/). – Здесь и далее прим. авт. 2 Большая часть информации в этой главе взята из двух обзоров: Montmerle, Augereau, Chaussidon, Gounelle, Marty, Morbidelli, 2006. Earth, Moon and Planets 98, doi: 10.1007/s11038-006-9087-5; Crida, 2009, http: //arxiv.org/abs/0903.3008. Другие источники указаны в тексте. 3 В последнее время возникли сомнения в биологическом происхождении графита в осадках Исуа. 4 Изомерами в химии называются вещества с одинаковым составом, но разным расположением атомов в молекулах. 5 Строго говоря, помимо этих трех путей фиксации СО2 существуют и другие: гидроксипропионатный, гидроксибутиратный и дикарбоксилатно-гидроксибутиратный циклы. По данным Браакмана и Смита, все они возникли в эволюции на основе восстановительного цикла Кребса уже после появления основных групп микроорганизмов, поэтому мы здесь их не рассматриваем. 6 Строго говоря, если мы найдем химическую систему, эволюционирующую не по Дарвину, а по Ламарку, то ее тоже следует считать живой. 7 Строго говоря, две цепи в ДНК антипараллельны (направлены навстречу друг другу), но нет оснований считать, что это обязательное требование к генетическим полимерам. 8 Диполем называют систему из двух электрических зарядов равной величины, но разных знаков, находящихся на малом расстоянии друг от друга. 9 Бактериофагами называются вирусы, заражающие бактерий. 10 Названия всех типов ферментов заканчиваются на «-аза». Название фермента обычно состоит из названий веществ, с которыми он работает, и реакции, которую он проводит. Например, аминоацил-тРНК-синтетаза соединяет аминокислоту и транспортную РНК. Алкоголь-НАДН-дегидрогеназа окисляет спирт, перенося два атома водорода с него на НАД. Эстеразы – это класс ферментов, расщепляющих сложноэфирные (C-O-CO) связи. Липазы – подкласс эстераз, расщепляющие сложноэфирную связь между глицерином и жирными кислотами в составе липидов (жиров). 11 Буквой N в последовательностях ДНК и РНК обозначают любой нуклеотид из четырех. Запись CGN обозначает сразу четыре кодона – CGU, CGC, CGA, CGG. 12 Маленькие молекулы ДНК, существующие в клетках наряду с основной молекулой ДНК – хромосомой. 13 Самые известные вещества из группы терпенов входят в состав эфирных масел растений и применяются в парфюмерии. Например, терпеновый спирт гераниол, из двух молекул которого собирается геранилгераниол, входит в масла герани, розы и лемонграсса. Однако терпены есть не только у растений, но и в любых других организмах – от бактерий до животных и человека. 14 У галоархей, населяющих пересоленые лагуны, есть бактериородопсин, и они могут использовать энергию света для получения АТФ. Но галоархеи – эволюционно молодая группа, получившая бактериородопсин горизонтальным переносом от бактерий. 15 В последнее время биологическое происхождение графита в осадках Исуа оспаривается, поэтому самые древние достоверные следы жизни имеют возраст 3, 5 млрд лет. 16 Пигментами в химии и биологии называются любые окрашенные вещества. 17 Эта глава, самая сложная во всей книге, расширяет и углубляет темы, рассмотренные в 15-й и 16-й главах. Если она показалась вам слишком сложной, не беспокойтесь, можете ее пропустить без вреда для восприятия книги в целом. 18 Клостридии – группа бактерий, из которой наиболее известны возбудители столбняка и ботулизма. Все клостридии живут в бескислородных условиях и получают энергию либо путем ацетогенеза, либо разными формами брожения. 19 Антипортерами называются транспортные белки, которые переносят два разных вещества навстречу друг другу. Натрий-протонный антипортер обменивает протон на ион натрия. 20 Метилотрофами называются микробы, которые способны питаться метаном и метиловым спиртом. О них рассказывалось в главе 11. 21 Мимивирусы, а также мегавирусы, пандоравирусы и питовирусы – недавно открытые гигантские ДНК-вирусы, паразитирующие на крупных амебах. Их вирусные частицы имеют размер до 1, 5 мкм и, в отличие от большинства вирусов, видны в световой микроскоп, а размер генома достигает 2, 5 млн пар нуклеотидов, что сравнимо с бактериями.
Предоставлено vk.com/nevzorovslibrary
|
Последнее изменение этой страницы: 2019-06-19; Просмотров: 86; Нарушение авторского права страницы