
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Базовая модель взаимодействия. Конкуренция. Отбор
Биологические системы вступают во взаимодействие друг с другом на всех уровнях, будь то взаимодействие биомакромолекул в процессе биохимических реакций, или взаимодействие видов в популяциях. Взаимодействие может протекать в структурах, тогда система может быть охарактеризована определенным набором состояний, так происходит на уровне субклеточных, клеточных и организменных структур. Кинетика процессов в структурах в математических моделях как правило описывается с помощью систем уравнений для вероятностей состояний комплексов. В случае, когда взаимодействие происходит случайно, его интенсивность определяется концентрацией взаимодействующих компонентов и их подвижностью обобщенной диффузией. Именно такие представления приняты в базовых моделях взаимодействия видов. Классической книгой, в которой рассматриваются математические модели взаимодействия видов стала книга Вито Вольтерра " Математическая теория борьбы за существование" (1931) [1]. Книга, построена как математический трактат, в ней постулированы в математической форме свойства биологических объектов и их взаимодействий, а затем эти взаимодействия исследуются как математические объекты, Именно с этой работы В. Вольтерра начались современная математическая биология и математическая экология. Вито Вольтерра (1860-1940) завоевал мировую известность своими работами в области интегральных уравнений и функционального анализа. Кроме чистой математики его всегда интересовали вопросы применения математических методов в биологии, физике, социальных науках. В годы службы в ВВС в Италии, он много работал над вопросами военной техники и технологии (задачи баллистики, бомбометания, эхо локации). В этом человеке сочетался талант ученого и темперамент активного политика, принципиального противника фашизма. Он был единственным итальянским сенатором, проголосовавшим против передачи власти Муссолини. Когда в годы фашистской диктатуры в Италии Вольтерра работал во Франции, Муссолини, желая привлечь на свою сторону всемирно известного ученого, предлагал ему различные высокие посты в фашистской Италии, но всегда получал решительный отказ. Антифашистская позиция привела Вольтерра к отказу от кафедры в Римском университете и от членства в итальянских научных обществах. Серьезно вопросами динамики популяций В. Вольтерра стал интересоваться с 1925 г. после бесед с молодым зоологом Умберто Д'Анкона, будущим мужем его дочери, Луизы. Д'Анкона, изучая статистику рыбных рынков на Адриатике, установил любопытный факт: когда в годы первой мировой войны (и сразу вслед за ней) интенсивность промысла резко сократилась, то в улове увеличилась относительная доля хищных рыб. Такой эффект предсказывался моделью " хищник-жертва", предложенной Вольтерра. Эту модель мы рассмотрим ниже. По сути дела это был первый успех математической биологии. Вольтерра предположил по аналогии со статистической физикой, что интенсивность взаимодействия пропорциональна вероятности встречи (вероятности столкновения молекул), то есть произведению концентраций. Это и некоторые другие предположения (См. Популяционная динамика ) позволили построить математическую теорию взаимодействия популяций одного трофического уровня (конкуренция) или разных трофических уровней (хищник-жертва). Системы, изученные Вольтерра, состоят из нескольких биологических видов и запаса пищи, который используют некоторые из рассматриваемых видов. О компонентах системы формулируются следующие допущения. . Пища либо имеется в неограниченном количестве, либо ее поступление с течением времени жестко регламентировано. . Особи каждого вида отмирают так, что в единицу времени погибает постоянная доля существующих особей. . Хищные виды поедают жертвы, причем в единицу времени количество съеденных жертв всегда пропорционально вероятности встречи особей этих двух видов, т.е. произведению количества хищников на количество жертв. . Если имеются пища в неограниченном количестве и несколько видов, которые способны ее потреблять, то доля пищи, потребляемая каждым видом в единицу времени, пропорциональна количеству особей этого вида, взятого с некоторым коэффициентом, зависящим от вида (модели межвидовой конкуренции). . Если вид питается пищей, имеющейся в неограниченном количестве, прирост численности вида за единицу времени пропорционален численности вида. . Если вид питается пищей, имеющейся в ограниченном количестве, то его размножение регулируется скоростью потребления пищи, т.е. за единицу времени прирост пропорционален количеству съеденной пищи. Перечисленные гипотезы позволяют описывать сложные живые системы при помощи систем обыкновенных дифференциальных уравнений, в правых частях которых имеются суммы линейных и билинейных членов. Как известно, такими уравнениями описываются и системы химических реакций. Действительно, согласно гипотезам Вольтерра, скорость процесса отмирания каждого вида пропорциональна численности вида. В химической кинетике это соответствует мономолекулярной реакции распада некоторого вещества, а в математической модели - отрицательным линейным членам в правых частях уравнений. Согласно представлениям химической кинетики, скорость бимолекулярной реакции взаимодействия двух веществ пропорциональна вероятности столкновения этих веществ, т.е. произведению их концентрации. Точно так же, согласно гипотезам Вольтерра, скорость размножения хищников (гибели жертв) пропорциональна вероятности встреч особей хищника и жертвы, т.е. произведению их численностей. И в том и в другом случае в модельной системе появляются билинейные члены в правых частях соответствующих уравнений. Наконец, линейные положительные члены в правых частях уравнений Вольтерра, отвечающие росту популяций в неограниченных условиях, соответствуют автокаталитическим членам химических реакций. Такое сходство уравнений в химических и экологических моделях позволяет применить для математического моделирования кинетики популяций те же методы исследований, что и для систем химических реакций. Можно показать, что вольтеровские уравнения могут быть получены не только из локального " принципа встреч", ведущего свое происхождение из статистической физики, но и исходя из баланса масс каждого из компонентов ценоза и энергетических потоков между этими компонентами. Рассмотрим простейшую из Вольтерра моделей модель отбора на основе конкурентных отношений. Эта модель работает при рассмотрении конкурентных взаимодействий любой природы биохимических соединений различного типа оптической активности, конкурирующих клеток, особей, популяций. Ее модификации применяются для описания конкуренции в экономике. Пусть имеется два совершенно одинаковых вида с одинаковой скоростью размножения, которые являются антагонистами, то есть при встрече они угнетают друг друга. Модель их взаимодействия может быть записана в виде:
Согласно такой модели, симметричное состояния сосуществования обоих видов является неустойчивым, один из взаимодействующих видов обязательно вымрет, а другой размножится до бесконечности. Введение ограничения на субстрат (типа 5) или системного фактора, ограничивающего численность каждого из видов (типа 2) позволяет построить модели, в которых один из видов выживает и достигает определенной стабильной численности. Они описывают известный в экспериментальной экологии принцип конкуренции Гаузе, в соответствии с которым в каждой экологической нише выживает только один вид. В случае, когда виды обладают различной собственной скоростью роста, коэффициенты при автокаталитических членах в правых частях уравнений будут различными, а фазовый портрет системы становится несимметричным. При различных соотношениях параметров в такой системе возможно как выживание одного из двух видов и вымирание второго (если взаимное угнетение более интенсивно, чем само регуляция численности), так и сосуществование обоих видов, в случае, когда взаимное угнетение меньше, чем самоограничение численности каждого из видов.
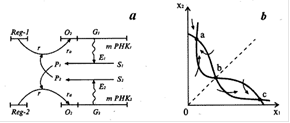
Рис.4. Схема синтеза двух ферментов Жакоба и Моно (а) и фазовый портрет триггерной систем (б) [6]. Еще одной классической триггерной системой является модель альтернативного синтеза двух ферментов Жакоба и Моно. Схема синтеза приведена на рис. 4а. Ген-регулятор каждой системы синтезирует неактивный репрессор. Этот репрессор, соединяясь с продуктом противоположной системы синтеза ферментов, образует активный комплекс. Активный комплекс, обратимо реагируя с участком структурного гена опероном, блокирует синтез mРНК. Таким образом, продукт второй системы Р2 является корепрессором первой системы, а Р1 - корепрессором второй. При этом в процессе корепрессии могут участвовать одна, две и более молекул продукта. Очевидно, что при таком характере взаимодействий при интенсивной работе первой системы вторая будет заблокирована, и наоборот. Модель такой системы предложена и подробно изучены в школе проф. Д.С.Чернавского [4] После соответствующих упрощений, уравнения, описывающие синтез продуктов Р1 и Р2 имеют вид:
Здесь P1, P2 - концентрации продуктов, величины A1, A2, B1, B2, выражаются через параметры своих систем. Показатель степени m показывает, сколько молекул активного репрессора (соединений молекул продукта с молекулами неактивного репрессора, который предполагается в избытке) соединяются с опероном для блокировки синтеза mRNK. Фазовый портрет системы, (изображение траекторий системы при разных начальных условиях на координатной плоскости, по осям которой отложены величины переменных системы), для m=2 изображен на рис.4б. Он имеет тот же вид, что и фазовый портрет системы двух конкурирующих видов. Сходство свидетельствует о том, что в основе способности системы к переключениям лежит конкуренция - видов, ферментов, состояний.
Рис.5. Модель химических реакций Лотки. Фазовый портрет системы при значениях параметров, соответствующих затухающим колебаниям.
|
Последнее изменение этой страницы: 2020-02-16; Просмотров: 153; Нарушение авторского права страницы

 (7)
(7)

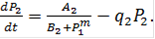 (8)
(8)