|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Трофическая структура биоценоза
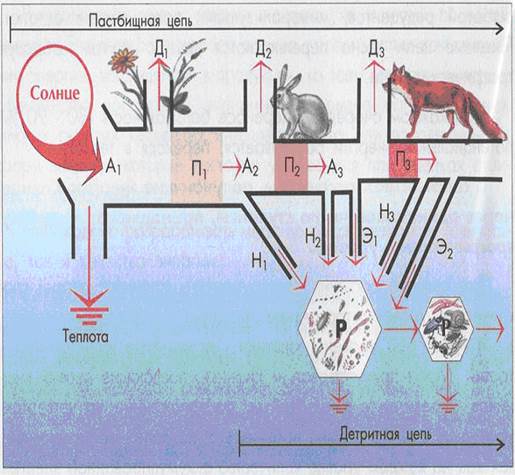
Каждый биоценоз, вследствие разной пищевой специализации составляющих еговидов, характеризуется определенной трофической структурой. Во всех биоценозах можно выделить два яруса: автотрофный и гетеротрофный. Автотрофы — это организмы, синтезирующие из неорганических соединений органические вещества с использованием энергии солнца или энергии, высвобождающейся при химических реакциях. К этой группе принадлежат все зеленые (хлорофиллсодержащие) растения, а также некоторые бактерии (пурпурные, железобактерии, серобактерии и др.). Гетеротрофы — организмы, использующие для своего питания готовые органические вещества, произведенные другими видами, и, как правило, не способные синтезировать вещества своего тела из неорганических составляющих. К гетеротрофам относятся все животные, паразитарные растения (заразиха, повилика), грибы и подавляющее большинство микроорганизмов. Пищевые цепи. Экологические пирамиды В экосистеме организмы связаны общностью энергии и питательных веществ, необходимых для поддержания жизни. Перенос вещества и энергии пищи, созданной растениями, через ряд организмов путем поедания одних видов другими называется цепью питания или пищевой (трофической) цепью. С термодинамической точки зрения живые организмы представляют собой преобразователей энергии. Главным источником энергии для большинства живых организмов является солнце. Используя его энергию, фотосинтезирующие организмы производят сложные органические вещества, в которых часть солнечной энергии запасена в виде энергии химических связей. Органические вещества служат источником пищи самим растениям и потребителям последних. Энергия, содержащаяся в пище, постепенно превращается в тепловую, рассеиваясь в окружающую среду. Каждое звено пищевой цепи называется трофическим уровнем. Организмы, получающие энергию солнца через одинаковое число звеньев пищевой цепи, принадлежат к одному трофическому уровню. I трофический уровень занимают продуценты – зеленые растения, которые в процессе фотосинтеза создают первичную биологическую продукцию. II уровень занимают консументы. Консументы питаются растениями (фитофаги) или другими гетеротрофами (зоофаги). В связи с этим различают консументов I порядка (фитофаги), II порядка (питаются фитофагами), IIIпорядка (хищники, питающиеся другими животными). III уровень занимают редуценты. Редуценты в процессе своей жизнедеятельности разрушают органические остатки, превращая их в неорганические соединения. К ним относятся бактерии, грибы, ряд животных, питающихся отмершими растениями и животными (дождевые черви, навозники, мертвоеды). Четко разделить все организмы биоценоза по трофическим уровням не всегда возможно, поскольку пищевая специализация позволяет некоторым видам занимать несколько трофических уровней. Выделяются два основных типа трофических цепей: пастбищные и детритные цепи. Первые начинаются уровнем продуцентов — зеленых растений. Начало детритной цепи дают растительные и животные остатки, экскременты. Энергия, заключенная в них, переходит к микроорганизмам (бактерии и грибы), разрушающим мертвое органическое вещество. Пастбищные трофические цепи, в свою очередь, можно разделить на пищевые цепи хищников и пищевые цепи паразитов. В цепи хищников соблюдается правило уменьшения числа особей, но увеличения их размеров. Если взять простую пастбищную цепь «злаки — мышевидные грызуны — лиса», то четко видно соблюдение данного правила. В той же цепи, но с участием паразитов, видно, что число последних возрастает, а их размеры существенно уменьшаются: «злаки — мышевидные грызуны — блохи — простейшие» или «злаки — корневые нематоды — бактерии». Детритные цепи начинаются с неживого органического вещества. К детритоядным организмам помимо бактерий и грибов относятся многие птицы (грифы, сипы, вороны), крабы, некоторые позвоночные и беспозвоночные (дождевые черви, жуки-навозники, могильщики, многие двукрылые, двупарноногие многоножки). Детритные цепи в наземных экосистемах часто имеют первостепенное значение. Они широко распространены в лесах, где очень много растительного опада, так как фитофаги используют здесь лишь около 10 % ежегодного прироста растительной массы. Для детритных цепей характерно усиленное повторное использование элементов питания, поскольку, высвобожденные одними организмами в виде экскрементов, они довольно быстро снова вовлекаются в оборот другими детритофагами. Пастбищные и детритные цепи в большинстве случаев находятся во взаимосвязи (см. рисунок 30).
Рисунок 30. Поток энергии в пастбищной и детринной пищевых цепях А1 – поглощенный растениями солнечный свет; А2 и А3 – корм, ассимилированный первичными и вторичными консументами; Д1, Д2, Д3 – траты энергии на дыхание; Н1, Н2, Н3 – неиспользованная часть корма; Э1, Э2 – экскременты; Р – редуценты; П1 – чистая первичная продукция; П2, П3 – вторичная продукция
Это вполне закономерно, ведь часть пищи, потребленной фитофагами, выделяется в виде неусвоенных остатков в окружающую среду. То же происходит и на других трофических уровнях. Вместе с тем, некоторые детритофаги могут сами являться пищей для хищников. Это и приводит к возникновению сложных взаимозависимостей между отдельными цепями и образованию трофических сетей в естественных биоценозах (см. рисунок 31).
Рисунок 31. Пищевая сеть
Структура и сложность сетей определяются типом биоценоза и набором организмов, образующих отдельные цепи. Перенос энергии с одного трофического уровня на другой никогда не бывает полным. Потери энергии связаны с тем, что: 1) не вся первичная продукция съедается, значительная ее часть отмирает, 2) не вся съеденная биомасса ассимилируется консументом, 3) не вся ассимилированная биомасса переходит в биомассу консумента. Эффективность пищевой цепи (экологическая эффективность) — это отношение ассимиляции на данном трофическом уровне к ассимиляции на предшествующем трофическом уровне. На уровне продуцентов это будет эффективность фотосинтеза. Экологическая эффективностьопределяется такими показателями, как эффективность потребления, эффективность ассимиляции и эффективность продуцирования. Эффективность потребления показывает, какая часть чистой продукции данного трофического уровня достается организмам следующего трофического уровня в виде пищи. Эффективность потребления продукции фитофагами в лесах в среднем составляет 5 %. В степи она почти в 5 раз выше, а в водных сообществах, где продуцентом является фитопланктон, достигает 50 %. Эффективность потребления у хищных позвоночных составляет более 50 % продукции, если жертвы — позвоночные и лишь 5 %, если беспозвоночные. Эффективность ассимиляции характеризует ту часть пищи, которая усваивается организмом и идет на его рост или для совершения работы. Эффективность ассимиляции низка у фитофагов, детритофагов и потребителей микроорганизмов (20—50 %). У хищников она достигает 80 %. Эффективность продуцирования — доля потребленной и усвоенной пищи, идущая на создание новой биомассы. Эффективность продуцирования у беспозвоночных составляет 30—40 %. У рыб, земноводных и других позвоночных организмов, температура тела которых зависит от температуры окружающей среды, эффективность продуцирования составляет 10 %. У теплокровных организмов, затрачивающих много энергии на поддержание температуры тела, она составляет 1—2 %. Закон (правило) Линдемана: потери энергии в процессах потребления пищи, ассимиляции и продуцирования, ее расход на дыхание приводят к тому, что на каждый последующий уровень поступает лишь около 10 % энергии предыдущего уровня.
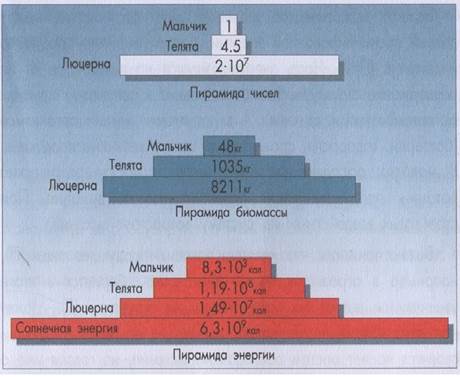
Правило Линдемана накладывает ограничения на длину пищевой цепи и численность организмов в ней участвующих. Ученые подсчитали, что среднее число трофических уровней для морского планктонного сообщества может равняться 7, для сообщества прибрежных вод — 5, степей — 4, влажных тропических лесов — 3. Соотношение между продуцентами, консументами и редуцентами в экосистеме, выраженное в их численности, биомассе или энергии и изображенное в виде графической модели, называется экологической пирамидой. Впервые экологическая пирамида была построена Ч. Элтоном (см. рисунок 32). Различают три типа экологических пирамид (см. рисунок 33): — пирамиды численности, отражающие численность отдельных организмов на каждом трофическом уровне; — пирамиды биомассы, отражающие количество органического вещества, синтезированного на каждом уровне; — пирамиды энергии, показывающие величину потока энергии на последовательных трофических уровнях. Число ступеней каждой пирамиды определяется числом трофических уровней, а ее форма сильно зависит от выбранного показателя. Наиболее просты пирамиды численности. Их форма различается для разных сообществ в зависимости от организмов-продуцентов. В лугово-пастбищном сообществе численность растений огромна, число их потребителей гораздо меньше, а количество хищников еще меньше. Вследствие этого пирамида будет иметь форму треугольника, резко убывающего к вершине. Если же построить пирамиду численности для леса, то ее форма будет иной. Уровень продуцентов здесь (на единицу площади) невелик. Поэтому основание пирамиды будет узким. Потребителей первого порядка (насекомые) будет значительно больше. Следовательно, второй блок пирамиды шире первого. В дальнейшем наблюдается уменьшение ширины блоков, и пирамида принимает вид, подобный лугово-пастбищному сообществу. Пирамиды биомассы для наземных сообществ обычно имеют ту же форму, что и пирамиды численности. В водных сообществах иногда наблюдаются «перевернутые» пирамиды. Интенсивное выедание фитопланктона снижает его биомассу, что, однако, не влияет на его продуктивность. Лучшее представление о сообществах дают пирамиды энергии. Они отражают картину скоростей прохождения массы пищи через пищевую цепь. Размер каждой ступеньки этой пирамиды определяется количеством энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за определенный период. Пирамиды энергии дают наиболее полное представление о функциональной организации сообщества. В отличие от пирамид численности и биомассы они всегда сужаются кверху при условии, что учтены все источники энергии, поступающей в экосистему с пищей.
Рисунок 32. Пирамида чисел Ч. Элтона (1927)
Рисунок 33. Пирамида чисел, биомасс и энергии, м-2∙ год-1 (по Ю. Одуму, 1975) Биогеоценоз (экосистема)
Живые организмы и их неживое окружение неразделимо связаны друг с другом и находятся в постоянном взаимодействии. Биогеоценоз — это однородный участок земной поверхности с определенным составом живых организмов (биоценоз) и условиями среды обитания (биотоп), объединенными обменом веществ и энергии в единый природный комплекс. Понятие «биогеоценоз» введено в науку В. Н. Сукачевым в 1940 г. Состав и строение биогеоценоза отражает рисунок 34.
Рисунок 34. Состав и взаимодействие компонентов биогеоценоза
Термин «биогеоценоз» очень близок (но не тождествен! ) к понятию «экосистема», предложенному в 1935 г. английским ботаником А. Тенсли и широко распространенному в зарубежных странах. Экосистема — это любая совокупность взаимодействующих живых организмов и условий среды, функционирующая как единое целое за счет обмена веществом, энергией и информацией с окружающей средой. Понятие «экосистема» не имеет ранга и размерности и может быть применимо к простым (сухое дерево), сложным (лес в целом) и искусственным (пруд) комплексам живых организмов. Биогеоценоз отличается от экосистемы определенностью своего объема. Его границы определены прежде всего характером растительного покрова или фитоценозом. Вследствие этого классификации экосистем довольно разнообразны. В зависимости от размера выделяют экосистемы: - микроэкосистема — экосистема упавшего дерева, пня, трупа животного и т. д.; - мезоэкосистема — экосистема луга, леса, озера; - макроэкосистема — экосистема суши, океана; - глобальная экосистема – биосфера. Широко используется классификация по биомам. Биом —это сочетание видов живого и окружающей их среды, составляющее экосистему географической зоны или сектора природного пояса. В частности, выделяют биомы тундры (арктической и альпийской), бореальных хвойных лесов, листопадных лесов, саванны, степей умеренной зоны, пустынь (травянистых и кустарниковых), тропических лесов. Среди водных экосистем выделяют экосистемы открытого океана, вод континентального шельфа, экосистемы заболоченных угодий, стоячих и текущих вод и т. д. Экосистема — основная функциональная единица живой природы, включающая организмы и абиотическую среду. Эти компоненты находятся во взаимодействии и необходимы для поддержания жизни на Земле.
Принцип формирования экосистемы: длительное существование организмов возможно лишь в рамках экосистем, где их компоненты и элементы дополняют друг друга и соответственно приспособлены друг другу. Принцип экологической комплементарности: никакая функциональная часть экосистемы не может существовать без других функционально дополняющих частей. Принцип экологической конгруэтности: функционально дополняя друг друга, живые составляющие экосистемы вырабатывают для этого соответствующие приспособления, скоординированные с условиями абиотической среды, в значительной мере преобразуемой теми же организмами. Закон внутреннего динамического равновесия: вещество, энергия, информация и динамические качества отдельных природных систем взаимосвязаны настолько, что любое изменение одного из этих показателей вызывает сопутствующие функционально-структурные количественные и качественные перемены, сохраняющие общую сумму вещественно-энергетических, информационных и динамических качеств систем, где эти изменения происходят. Следствия из закона внутреннего динамического равновесия: - любое изменение среды приводит к развитию цепных реакций, направленных на нейтрализацию изменения, - взаимодействие вещественно-энергетических и экологических компонентов, информации и динамических качеств природных систем количественно нелинейно, - изменения системы относительно необратимы, - любое местное преобразование в глобальном масштабе вызывает реакции, приводящие к относительной неизменности эколого-экономического потенциала.
Свойства и закономерности развития экосистем: Аксиома эмерджентности: целое всегда имеет особые свойства, отсутствующие у отдельных частей, целое не равно механической сумме элементов. Закон необходимого разнообразия: система не может состоять из абсолютно одинаковых элементов. Закон полноты составляющих: число функциональных составляющих системы и связей между ними должно быть оптимальным – без недостатка или избытка. Закон развития системы за счет окружающей ее среды: любая система может развиваться только за счет использования материально-технических и информационных возможностей окружающей ее среды, абсолютно изолированное саморазвитие невозможно. Т.е., экосистемы — это системы открытые с термодинамической точки зрения. Правило системно-динамической комплементарности: любая саморазвивающаяся система состоит из двух видов структур, один из которых сохраняет и закрепляет ее строение и функциональные особенности, а другой способствует видоизменению системы с образованием новой функционально-морфологической специфики. Правило основного обмена: любая большая динамическая система в стационарном состоянии использует приход энергии, вещества и информации, главным образом, для своего самоподдержания и саморазвития. Теорема сохранения упорядоченности: в открытых системах энтропия падает до минимальной, но большей 0, величины. При этом вещество в системе распределено неравномерно. Закон минимума диссипации энергии: при вероятности развития процесса в некотором множестве направлений, допускаемых законами термодинамики, реализуется то, которое обеспечивает минимум рассеивания энергии (минимум роста энтропии). Закон максимизации энергии и информации: наилучшими шансами на самосохранение обладает система, в наибольшей степени способствующая поступлению, выработке и эффективному использованию энергии и информации.
Способность к саморегулированию и самоподдержанию экосистемы называется гомеостазом. В его основе лежит принцип отрицательной обратной связи, уменьшающей отклонение системы от состояния равновесия. Благодаря этому принципу регулируются процессы запасания и высвобождения питательных веществ, продуцирования и разложения органических соединений. Относительную стабильность экосистем обеспечивают устойчивый круговорот веществ и поток энергии.
Продуктивность экосистем
Каждая экосистема характеризуется способностью к самовозобновлению, т. е. созданию (продуцированию) новой биомассы. Биомасса — выраженное в единицах массы или энергии количество живого вещества тех или иных организмов, приходящееся на единицу площади или объема. Воспроизведение биомассы растений, микроорганизмов и животных, входящих в состав биоценоза, называется биологической продуктивностью. Она представляет собой количество воспроизведенной биомассы на 1 м2 площади (1 м3 объема) в единицу времени и выражается в граммах углерода или сухого вещества. Скорость создания органического вещества в любой экосистеме называется биологической продукцией. Различают продукцию первичную и вторичную. Первичная продукция — скорость образования биомассы первичными продуцентами (растениями). Она может быть валовой и чистой. Валовая первичная продукция (ВПП) — скорость, с которой растения накапливают энергию в виде химических связей (в процессе фотосинтеза). Часть этой энергии расходуется в процессе жизнедеятельности, т. е. тратится на дыхание (Д). Поэтому чистая первичная продукция (ЧПП) представляет собой фактическую скорость накопления органического вещества с учетом затрат на дыхание:
ЧПП = ВПП -Д
Чистая первичная продукция сильно зависит от типа экосистемы. Наиболее велики ее значения для влажных тропических лесов, зарослей водорослей и рифов, болот и маршей (2000—2500 г/м2 в год). В листопадных лесах и тайге она составляет 800—1200 г/м2 в год, степях и саваннах — 600—900 г/м2 в год. Очень низкими значениями чистой первичной продукции характеризуются тундра, высокогорья, пустыни и полупустыни (порядка 90—140 г/м2 в год). Среди водных экосистем велика продуктивность в речных дельтах (1500 г/м2 в год). Чистая первичная продукция материковых экосистем в 5 раз выше (773 г/м2 в год), чем морских (152 г/м2 в год). На культивируемых землях, где производство биомассы поддерживается человеком, чистая первичная продукция достигает 150 г/м2 в год. В целом же для биосферы этот показатель в среднем составляет около 333 г/м2 в год. Скорость продуцирования биомассы гетеротрофами называется вторичной продукцией. Энергетический баланс на уровне консументов включает в себя затраты на рост и дыхание, а также потери энергии с неусвоенной и выделенной в виде экскрементов пищей. Из этих составляющих лишь энергия, используемая на дыхание, рассеивается в окружающую среду. Энергия, заключенная в экскрементах, передается редуцентам и детритофагам. Доля чистой первичной продукции, переходящей в детритные цепи редуцентов, в разных экосистемах неодинакова. Она выше в лесах, где около 90 % первичной продукции не потребляется в пастбищных цепях. Поскольку при переходе от одного трофического уровня на другой происходят потери энергии, то продукция каждого последующего уровня меньше продукции предыдущего. Продукция, которая выводится за пределы биоценоза, называется конечной . Ее получает человек, например на сельхозугодьях, а также в процессе охоты и рыболовства.
Развитие экосистем
Ни один биогеоценоз не существует неограниченно долгое время. В процессе жизнедеятельности организмов изменяются условия среды их обитания. Длительное существование популяций может так изменить биотоп, что он становится малопригодным для одних видов, но благоприятным для других. В результате на данном месте развивается другой, более приспособленный к новым условиям биоценоз. Популярное: |
Последнее изменение этой страницы: 2016-03-25; Просмотров: 3521; Нарушение авторского права страницы