|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
После введения раствора Рингера,
После введения арэн-терина. Рис. 8.3, Торможение потребления глюкозы после внутрибрюшинного введения препаратов арэнтерина, секретина и холецистокинина (по: Уголев, 1978). По оси абсцисс - время после потребления глюкозы (мин); по оси ординат - торможение потребления 40%-ного раствора глюкозы (% по отношению к исходному уровню). 1 - после введения арэнтерина; 2 - после введения секретина; После введения холецистокинина. _____________________________
жения аппетита и возникновения физиологической жажды. С этой точки зрения, можно думать, что в регуляции аппетита опосредованно участвуют: секретин, который стимулирует секрецию жидкой части поджелудочного сока, и гастрин, который вызывает значительную по объему секрецию желудочного сока. Кроме того, растяжение желудка, ацидификация желудка и верхних отделов двенадцатиперстной кишки являются также источником нервных сигналов, передаваемых главным образом по блуждающим нервам. Особого рассмотрения заслуживает роль кишечной гормональной системы в потреблении пищи в связи с
термостатической теорией регуляции аппетита. Ряд данных позволяет считать, что СДД пищи и изменение внутренней, в частности гипоталамической, температуры могут быть важным компонентом нормального насыщения. Так, показано, что повреждения ростральной области гипоталамуса, связанной с регуляцией как температуры, так и потребления пищи, приводят к нарушению обеих этих функций (обзор: Hamilton, 1967). Здесь необходимо напомнить о существовании обнаруженных нами компонентов СДД пищи (см. гл. 7), контролируемых кишечной гормональной системой и действием желудочного химуса при его прохождении через двенадцатиперстную кишку. Понижение пищевого возбуждения, связанное с интенсификацией обмена, и повышение внутренней температуры также в определенной мере обусловлены включением кишечного гормонального комплекса. В целом прохождение пиши через верхние отделы пищеварительного аппарата вызывает выделение ряда гормонов желудочно-кишечного тракта, прямо или косвенно влияющих на переход от голодного состояния к сытому и во многих случаях к появлению питьевых реакций. К гормонам, вызывающим физиологическую дегидратацию, которая благоприятствует понижению пищевого возбуждения и повышению питьевого, относятся гастрин и секретин. Дегидратация может влиять на пищевой центр как через периферические, так и через центральные осморецепто-ры. В свете обсуждаемых данных обращает на себя внимание недавнее исследование, на основании которого высказано предположение, что мотилин играет важную роль в контроле моторной деятельности не только у собак и людей, но и у свиней (Bull, Sis-sons, 1988). Совершенная регуляция потребления пищи включает в себя два типа регуляторов, различающихся по своим временным характеристикам: быстродействующие (кратковременные) и длительнодействующие (долговременные). Как правило, регуляция первого типа обеспечивает срочные, но не вполне точные реакции, тогда как медленные регуляции обеспечивают точное соответствие между потребностью в калориях и их поглощением. Таким образом, если существует несколько типов кишечных гормонов, влияющих на аппетит (а их должно быть несколько, в том числе арэнтерин и холецистокинин), то с их помощью реализуются как более быстрые, так и более медленные типы регуляции аппетита, или потребления пищи (рис. 8.4).
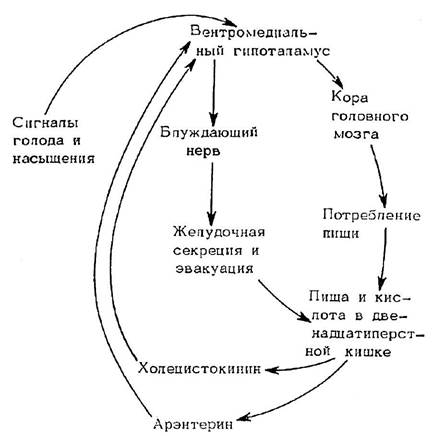
Рис. 8.4. Схема нейрогуморальной регуляции аппетита с участием арэнтерина (по: Уголев, 1978). _____________________________
До сих пор мы рассматривали влияние кишечной гормональной системы на пищевой центр, сузив эту проблему до регуляции потребления пищевых веществ. В действительности речь идет о том, что кишечная гормональная система путем изменения мотиваций участвует в переключении целенаправленного поведения, характерного для голодного животного, на поведение, свойственное сытому животному. Поэтому можно думать, что чем дальше будет развиваться эта проблема, тем больше места в ней будет занимать область нейротропных и психотропных влияний кишечной гормональной системы. Как отмечено выше, Н. Дафни е. Джекобсон (Dafny, Jacobson, 1975) показали, что гормоны желудочно-кишечного тракта, продуцируемые при потреблении пищи, индуцируют насыщение посредством изменения электрической активности центральной нервной системы. По мнению авторов, центр аппетита взаимодействует с вышележащими центрами, которые могут влиять на тонус сосудов, моторику пищевари-
тельного аппарата и секреторную активность. Предполагается, что холецистокинин может действовать как модулятор одного из известных химических мессенджеров: Вместе с тем эти данные подтверждают высказанную нами мысль, что эффекты гормонов желудочно-кишечного тракта должны охватывать в конце концов более значительную сферу реакций, чем только контроль над пищеварительными процессами. В целом приведенные сведения свидетельствуют, что кишечная гормональная система может играть важную роль не только в контроле обмена веществ за пределами пищеварительного аппарата, но и в управлении поведенческими реакциями, прямо или косвенно связанными с регуляцией потребления пищи.
Заключительные замечания (точность регуляции потребления пищи и две теории питания)
Концепция идеальной регуляции потребления пищи, по-видимому, соответствует концепции идеального питания. В действительности организмы не являются идеальными системами (см. подробно обзоры: Уголев, 1985, 1987а), и регуляция потребления пищи происходит с большой степенью приближенности. При некоторых режимах питания, например при потреблении высококалорийной пищи, может происходить переедание, а при переходе к низкокалорийной - в течение некоторого времени недоедание. За непродолжительные интервалы времени возникают нарушения баланса (деэквилибрация) ряда компонентов внутренней среды как у животных (в естественных условиях их существования), так и у человека (в значительной мере в искусственных условиях жизни). Однако после этого происходит восстановление нарушенного баланса (реэквилибрация) в результате интегрального эффекта изменения выбора пищи и вегетативной регуляции гомеостаза. В нашей лаборатории показано, что солевой баланс у крыс на оптимальном уровне поддерживается не в каждый данный момент, а как результат взаимодействия различных регуляторных систем на протяжении продолжительного времени. Каким же образом, несмотря на неидеальное управление потреблением пищи, поддерживается высокий уровень жизнедеятельности? (Неидеальное управление питанием понимается в том смысле, что в каждый данный момент нет полного соответствия между потерей и поступлением пищевых веществ и, следователь- но, нет сохранения молекулярного состава организма.) По всей вероятности, гомеостаз внутренних сред существует, при этом в случае избыточного поглощения пиши большую роль играет депонирование избытка веществ, а в случае недостаточного поглощения - мобилизация депо, точно так же как и изменение уровня функционирования ряда систем. Такие представления о регуляции потребления пищи больше соответствуют теории адекватного питания с ее многочисленными видовыми и экологическими вариациями, чем концепции идеального питания в свете теории сбалансированного питания. Возникает вопрос, как происходит узнавание недостающих веществ, учитывая множество незаменимых элементов, при том, что каждый из них должен быть опознан? Можно выдвинуть гипотезу, согласно которой существуют опознающие рекогносцирующие клетки или органы (например, у высших организмов язык), которые в условиях деэквилибрации узнают недостающий во внутренней среде элемент, если он содержится в пище. Например, если для нормального функционирования системы необходимы элементы А, Б и В, то в отсутствие элемента В система работает в измененном режиме. Но при контакте с пищей, содержащей недостающий элемент В, работа системы нормализуется и, следовательно, система специфически опознает такой элемент. Что касается физиологической роли различных рецепторных зон, то в настоящее время она не может быть полностью охарактеризована. Но уже сегодня можно говорить, что разные рецепторные зоны контролируют различные процессы. Например, нами было обнаружено, что при субдиафрагмальной ваготомии суточное потребление калорий не меняется, но меняется ритм потребления пищи; при выключении синокаротидной рефлексогенной зоны меняется уровень потребления пищи и воды. Наконец, имеются серьезные основания полагать, что наряду с классическими гормонами желудочно-кишечного тракта существуют гипоталамотропные кишечные -гормоны, среди которых, в частности, наиболее подробно изучен аппетитрегулирующий гормон - арэн-терин. Этот гормон, вызывая чувство сытости, тормозит потребление пищи в значительно большей мере, чем холецистокинин. Состояние сытости, вызываемое холецистокинином, реализуется главным образом через блуждающие нервы. Эффект холецистокинина может быть предупрежден субдиафрагмальной ваготомией. С другой стороны, насыщающий эффект арэнтерина не снижается после субдиафрагмальной ваготомии, что говорит о прямом центральном действии этого гормона. Таким образом, кишечная гормональная система играет существенную роль в регуляции аппетита. Большой интерес представляют данные о взаимодействии гормональных и рефлекторных сигналов. Влияние различных диет на симпатическую нервную систему представлено в недавнем обзоре (Landsberg, Young, 1985).
Глава 9 Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 649; Нарушение авторского права страницы