
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Н.К.Кольцов. Наследственные молекулы
*** Первоначально, когда у простейших организмов впервые слагались генонемные молекулы, они были представлены однообразными более или менее длинными цепями одинаковых звеньев, вроде кератина или серицина. Каждое звено состояло из немногих простых радикалов. При дальнейшей эволюции организма эти молекулы постепенно усложнялись путем присоединения к некоторым звеньям боковых радикалов, получающих значение генов. Мало-помалу число этих боковых цепей, размещенных в определенных пунктах генонемы, увеличивалось, и самые радикалы все более усложнялись. Микроскопическая длина хромосом в слюнных железах дрозофилы представляет картину уже очень высокой дифференцировки генонем. Если признать, что поперечные диски соответствуют генам, то здесь мы должны поместить именно боковые радикалы или цепи радикалов, которые адсорбируют ярко окрашенный хроматин. В таком случае неокрашиваемые сегменты, в которых мы различаем продольные нити, придется признать основными цепями, не осложненными сложными боковыми придатками. Но при дальнейшей дифференцировке и сюда могут присоединиться боковые радикалы - новые гены, а с другой стороны, уже имеющиеся боковые радикалы могут осложняться упрощаться в мутационном процессе. *** Мы, конечно, не должны увлекаться достигнутыми успехами, тем более что в своей химической части они далеки от завершения, более того - еще весьма спорны. За нашей нынешней синтезой еще придет новая антитеза, но это будет уже новый этап развития науки. И вряд ли, по крайней мере у нас в Советском Союзе, найдется хотя бы один ученый, который решился бы объявить все эти научные изыскания бесплодными и никчемными. Кольцов Н. К. Наследственные молекулы. – Л., 1958. - С. 93-118. Т. Морган. Теория гена
Работы Менделя были вновь открыты в 1900 году. Четыре года спустя Бэтсон и Пеннет сообщили о наблюдениях, которые не давали тех числовых результатов, которые нужно было бы ожидать для двух независимых пар признаков. Например, если душистый горошек, имеющий цветы пурпурного цвета и удлиненные пыльцевые зерна, скрещивается с горошком с красными цветами и круглыми пыльцевыми зернами, то те два признака, которые наследуются совместно от одного из родителей, появляются вместе у потомства чаще, чем следовало бы ожидать при независимом распределении признаков... Бэтсон и Пеннет считали, что эти результаты являются следствием «отталкивания», между комбинациями пурпурного - длинного и красного - круглого, происходящими от противоположных родителей. В настоящее время явление это называется как раз наоборот - сцеплением. Под этим термином мы подразумеваем тот факт, что когда некоторые признаки вступают в скрещивание совместно, они проявляют тенденцию оставаться вместе и у последующих поколений или, выражаясь иначе, некоторые пары признаков не свободны в своем распределении. Таким образом, поскольку сцепление имеет место, оказывается, что разделение наследственного вещества является до некоторой степени ограниченным. Например, у плодовой мухи Drosophila melanogaster известно около 400 новых типов мутантов, особенности которых составляют всего лишь четыре группы сцепления. *** Члены группы сцепления могут иногда оказаться не так полно сцепленными друг с другом, ... некоторые из рецессивных признаков одной серии могут оказаться замененными признаками дикого типа из другой серии. Однако даже и в этом случае они все-таки считаются сцепленными, потому что соединенными вместе они остаются чаще, чем наблюдается такой обмен между сериями. Этот обмен называется перекрестом (crossing-over). Термин этот обозначает, что между двумя соответственными сериями сцепления может происходить правильный обмен их частями, в котором участвует большое число генов. *** Теперь мы можем сформулировать теорию гена. Эта теория устанавливает, что признаки или свойства особи являются функцией соединенных в пары элементов (генов), заложенных в наследственном веществе в виде определенного числа групп сцепления; она устанавливает затем, что члены каждой пары генов, когда половые клетки созревают, разделяются в соответствии с первым законом Менделя и, следовательно, каждая зрелая половая клетка содержит только один ассортимент их; она устанавливает также, что члены, принадлежащие к различным группам сцепления, распределяются при наследовании независимо, соответственно второму закону Менделя; равным образом она устанавливает, что иногда имеет место закономерный взаимообмен - перекрест - между соответственными друг другу элементами двух групп сцепления; наконец, она устанавливает, что частота перекреста доставляет данные, доказывающие линейное расположение элементов по отношению друг к другу. Эти принципы, которые я беру на себя смелость, объединив их вместе, назвать теорией гена, дают нам возможность поставить изучение проблем генетики на строго числовом основании и позволяют предсказывать с большой степенью точности, что произойдет в том или другом случае. Морган. Теория гена. – Л., 1927. – С. 132-136, 174-180. Популярное:
|
Последнее изменение этой страницы: 2016-05-29; Просмотров: 1006; Нарушение авторского права страницы
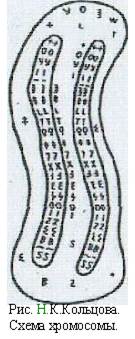 Свою гипотезу молекулярного строения хромосомы я могу иллюстрировать схемой, опубликованной мной в 1928 г. На рисунке изображена хромосома, внутри которой проходят две генонемы, как это бывает обыкновенно задолго перед делением клетки. Каждая генонема представляет собой пучок длинных молекул, из которых на рисунке изображены только две. Все четыре молекулы имеют совершенно одинаковое строение и состоят из ряда белковых радикалов, связанных между собой главными валентностями. В каждом пучке сходные молекулы сдерживаются боковыми связями. Большая часть хромосомы между генонемами и оболочкой хромосомы (хромолеммой) заполнена хромоплазмой и хроматином, в состав которого в качестве элемента обмена веществ входят те же самые радикалы — гены, из которых состоит генонема, или частицы, обломки этих радикалов, а также нуклеиновая кислота. При росте генонемного пучка молекул эти радикалы располагаются так же, как при кристаллизации, именно в тех местах кристаллической решетки, где находятся такие же радикалы. На схеме с внутренней стороны генонем нарисовано несколько уже сложившихся отрезков. Когда толщина генонемного пучка молекул путем обрастания доходит до известного предела, генонема расщепляется вдоль. В разные моменты жизни клетки обмен радикалами может идти в разные стороны: то из нуклеоплазмы в хромосому, то из хромосомы в нуклеоплазму.
Свою гипотезу молекулярного строения хромосомы я могу иллюстрировать схемой, опубликованной мной в 1928 г. На рисунке изображена хромосома, внутри которой проходят две генонемы, как это бывает обыкновенно задолго перед делением клетки. Каждая генонема представляет собой пучок длинных молекул, из которых на рисунке изображены только две. Все четыре молекулы имеют совершенно одинаковое строение и состоят из ряда белковых радикалов, связанных между собой главными валентностями. В каждом пучке сходные молекулы сдерживаются боковыми связями. Большая часть хромосомы между генонемами и оболочкой хромосомы (хромолеммой) заполнена хромоплазмой и хроматином, в состав которого в качестве элемента обмена веществ входят те же самые радикалы — гены, из которых состоит генонема, или частицы, обломки этих радикалов, а также нуклеиновая кислота. При росте генонемного пучка молекул эти радикалы располагаются так же, как при кристаллизации, именно в тех местах кристаллической решетки, где находятся такие же радикалы. На схеме с внутренней стороны генонем нарисовано несколько уже сложившихся отрезков. Когда толщина генонемного пучка молекул путем обрастания доходит до известного предела, генонема расщепляется вдоль. В разные моменты жизни клетки обмен радикалами может идти в разные стороны: то из нуклеоплазмы в хромосому, то из хромосомы в нуклеоплазму.