
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Структура и функционирование экосистем
Структура экосистем. С точки зрения трофической структуры экосистему можно разделить на два яруса – автотрофный и гетеротрофный (по Ю. Одуму, 1986). 1. Автотрофный ярус или «зеленый пояс» включающий растения или их части, содержащие хлорофилл, где преобладают фиксация энергии света, использование простых неорганических соединений и накопление сложных органических соединений. 2. Гетеротрофный ярус или «коричневый пояс» почв и осадков, разлагающихся веществ, корней и т.д., в котором преобладают использование, трансформация и разложение сложных соединений. По Ю. Одуму, с биологической точки зрения в составе экосистемы удобно выделить следующие компоненты: 1) неорганические вещества, 2) органические вещества, 3) воздушную, водную и субстратную среду, 4) продуцентов, 5) макроконсументов, 6) микроконсументов. 1. Неорганические вещества (СО2, Н2О, N2, О2 минеральные соли и др.), включающиеся в круговороты. 2. Органические вещества (белки, углеводы, липиды, гумусовые вещества и др.), связывающие биотическую и абиотическую части. 3. Воздушная, водная и субстратная среда, включающая абиотические факторы. 4. Продуценты – автотрофные организмы, способные производить органические вещества из неорганических, используя фотосинтез или хемосинтез (растения и автотрофные бактерии). 5. Консументы (макроконсументы, фаготрофы) – гетеротрофные организмы, потребляющие органическое вещество продуцентов или других консументов (животные, гетеротрофные растения, некоторые микроорганизмы). Консументы бывают первого порядка (фитофаги, сапрофаги), второго порядка (зоофаги, некрофаги) и т.д. 6. Редуценты (микроконсументы, деструкторы, сапротрофы, осмотрофы) – гетеротрофные организмы, питающиеся органическими остатками и разлагающие их до минеральных веществ (сапротрофные бактерии и грибы). Следует учитывать, что и продуценты, и консументы частично выполняют функции редуцентов, выделяя в окружающую среду минеральные вещества – продукты их метаболизма.
Пищевые цепи и сети. Питаясь друг другом, живые организмы образуют цепи питания. Цепь питания – последовательность организмов, по которой передается энергия, заключенная в пище, от ее первоначального источника. Каждое звено цепи называется трофическим уровнем. Первый трофический уровень – продуценты (автотрофные организмы, преимущественно зеленые растения). Второй трофический уровень – консументы первого порядка (растительноядные животные и паразиты продуцентов). Третий трофический уровень – консументы второго порядка (первичные хищники, питающиеся растительноядными животными, и паразиты первичных консументов). Четвертый трофический уровень – консументы третьего порядка (вторичные хищники, питающиеся плотоядными животными, и паразиты вторичных консументов). В пищевой цепи редко бывает больше 4–6 трофических уровней. Последний трофический уровень – редуценты (сапротрофные бактерии и грибы). Они осуществляют минерализацию – превращение органических остатков в неорганические вещества. Редуценты могут представлять любой трофический уровень, начиная со второго. Различают два типа пищевых цепей. Цепи выедания (или пастбищные) – пищевые цепи, начинающиеся с живых фотосинтезирующих организмов. Например, фитопланктон → зоопланктон → рыбы микрофаги → рыбы макрофаги → птицы ихтиофаги. Цепи разложения (или детритные) – пищевые цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных. Например, детрит → детритофаги → хищники микрофаги → хищники макрофаги. Таким образом, поток энергии, проходящий через экосистему, разбивается как бы на два основных направления. Энергия к консументам поступает через живые ткани растений или через запасы мертвого органического вещества. Цепи выедания преобладают в водных экосистемах, цепи разложения – в экосистемах суши. В сообществах пищевые цепи сложным образом переплетаются и образуют пищевые сети. В состав пищи каждого вида входит обычно не один, а несколько видов, каждый из которых в свою очередь может служить пищей нескольким видам. С одной стороны, каждый трофический уровень представлен многими популяциями разных видов, с другой стороны, многие популяции принадлежат сразу к нескольким трофическим уровням. В результате благодаря сложности пищевых связей выпадение какого-то одного вида часто не нарушает равновесия в экосистеме.
6.6. Круговорот веществ и поток энергии в экосистеме. Биологическая продуктивность экосистем Принцип единства организм-среда (основной биологический закон) – между живыми организмами и окружающей их средой существуют тесные взаимоотношения, взаимозависимости и взаимовлияния, обуславливающие их единство. В экосистеме органические вещества синтезируются автотрофами из неорганических веществ. Затем они потребляются гетеротрофами. Выделенные в процессе жизнедеятельности или после гибели организмов (как автотрофов, так и гетеротрофов) органические вещества подвергаются минерализации, то есть превращению в неорганические вещества. Эти неорганические вещества могут быть вновь использованы автотрофами для синтеза органических веществ. Так осуществляется биологический круговорот веществ. В то же время, энергия не может циркулировать в пределах экосистемы.Поток энергии (передача энергии), заключенной в пище, в экосистеме осуществляется однонаправленно от автотрофов к гетеротрофам. Жизнь на Земле продолжается, не прерываясь потому, что она протекает в системе биологических круговоротов вещества и поддерживается постоянным потоком солнечной энергии. По I закону термодинамики, энергия не исчезает бесследно, а переходит из одной формы в другую. Так, на первом трофическом уровне зелеными растениями солнечная энергия в процессе фотосинтеза преобразуется в энергию химических связей органических веществ. Это валовая первичная продукция . По II закону термодинамики, любые превращения энергии сопровождаются переходом части ее в такое состояние, когда она уже не может быть использована для работы. Так, большая часть поглощенной растениями, но не усвоенной энергии, рассеивается в окружающую среду в виде тепловой энергии. Часть образованных органических веществ окисляется, а высвобождающаяся энергия расходуется на поддержание всех метаболических процессов. Это так называемые траты на дыхание. Эта энергия, в конечном счете, также рассеивается в виде тепла. Оставшаяся часть новообразованных органических веществ составляет прирост биомассы растений и называется чистой первичной продукцией. В чистую первичную продукцию превращается только 1% поглощенной растением энергии. До второго трофического уровня доходит только часть чистой первичной продукции. Некоторая ее часть не используется консументами первого порядка. Она может накапливаться или экспортироваться за пределы системы. Та часть, которую ассимилировали (потребили) консументы, частично тратится на дыхание, частично выделяется с экскрементами, а остальное накапливается в виде вторичной продукции . Вторичная продукция на каждом последующем трофическом уровне консументов и т.д.) составляет около 10% предыдущей (хотя на уровне хищников может быть выше – около 20%). В результате, чем длиннее пищевая цепь, тем меньше остается к ее концу накопленной в органическом веществе энергии. Поэтому число трофических уровней никогда не бывает слишком большим. Таким образом, при передаче энергии с одного трофического уровня на другой большая часть энергии рассеивается в виде тепла (в соответствии со вторым законом термодинамики), и только около 10 % от первоначального количества передается по пищевой цепи. Правило десяти процентов – на каждый следующий трофический уровень переходит примерно 10 % вещества и энергии предыдущего уровня. Пищевые цепи можно представить в виде экологических пирамид. Различают три основных типа экологических пирамид. Пирамида чисел(пирамида Элтона) отражает уменьшение численности организмов от продуцентов к консументам. Пирамида биомасс показывает изменение биомасс на каждом следующем трофическом уровне: для наземных экосистем пирамида биомасс сужается кверху, для экосистемы океана – имеет перевернутый характер (сужается книзу), что связано с быстрым потреблением фитопланктона консументами. Пирамида энергии (продукции) имеет универсальный характер и отражает уменьшение количества энергии, содержащейся в продукции, создаваемой на каждом следующем трофическом уровне. Прирост биомассы в экосистеме, созданной за единицу времени, называется биологической продукцией (продуктивностью). Различают первичную и вторичную продукцию сообщества. Первичная продукция – биомасса, созданная за единицу времени продуцентами. Она делится на валовую и чистую. Валовая первичная продукция (общая ассимиляция) – это общая биомасса, созданная растениями в ходе фотосинтеза. Часть ее расходуется на поддержание жизнедеятельности растений – траты на дыхание (40–70 %). Оставшаяся часть составляет чистую первичную продукцию (чистая ассимиляция), которая в дальнейшем используется консументами и редуцентами, или накапливается в экосистеме. Вторичная продукция – биомасса, созданная за единицу времени консументами. Она различна для каждого следующего трофического уровня. Масса организмов определенной группы (продуцентов, консументов, редуцентов) или сообщества в целом называется биомассой. Самой высокой биомассой и продуктивностью обладают тропические дождевые леса, самой низкой – пустыни и тундры. Если в экосистеме скорость прироста растений (образования первичной продукции) выше темпов переработки ее консументами и редуцентами, то это ведет к увеличению биомассы продуцентов. Если при этом присутствует недостаточная утилизация продуктов спада в цепях разложения, то происходит накопление мертвого органического вещества. Это проявляется в заторфовывании болот, образовании мощной лесной подстилки и т.п. В стабильных экосистемах биомасса остается постоянной, так как практически вся продукция расходуется в цепях питания.
Динамика экосистем Изменения в сообществах могут быть циклическими и поступательными. I. Циклические изменения – периодические изменения в биоценозе (суточные, сезонные, многолетние), при которых биоценоз возвращается к исходному состоянию. Суточные циклы связаны с изменением освещенности, температуры, влажности и других экологических факторов в течение суток. Суточные ритмы проявляются в изменении состояния и активности живых организмов в течение суток. Например, чередование световой и темновой фаз фотосинтеза у растений, сна и бодрствования у животных. Сезонная цикличность связана с изменением экологических факторов в течение года и наиболее сильно выражена в высоких широтах, где велик контраст зимы и лета. Сезонная изменчивость проявляется не только в изменении состояния и активности, но и количественного соотношения отдельных видов. На определенный период многие виды выключаются из жизни сообщества, впадая в спячку, оцепенение, перекочевывая или улетая в другие районы. Многолетняя изменчивость связана с флуктуациями климата или другими внешними факторами (степень разлива рек), либо с внутренними причинами (особенности жизненного цикла растений-эдификаторов, повторения массового размножения животных). II. Поступательные изменения – изменения в биоценозе, в конечном счете приводящие к смене этого сообщества другим. Сукцессия – последовательная смена биоценозов (экосистем), выраженная в изменении видового состава и структуры сообщества. Последовательный ряд сменяющих друг друга в сукцессии сообществ называется сукцессионной серией. К сукцессиям относятся опустынивание степей, зарастание озер и образование болот и др. В основе возникновения сукцессий лежит неполнота круговорота веществ в сообществе. Если в биоценозах деятельность одних видов не компенсирует деятельность других, то условия среды неминуемо изменяются. Популяции меняют среду в неблагоприятную для себя сторону и вытесняются другими видами, для которых новые условия экологически более выгодны. Этот процесс продолжается до тех пор, пока не сформируется уравновешенное сообщество, которое способно поддержать баланс веществ в экосистеме. Так, можно проследить процесс зарастания зарастание небольших озер – из-за недостатка кислорода в придонных слоях организмы-разлагатели не в состоянии обеспечить полный распад отмирающих растений, образуются торфянистые отложения, озеро мелеет, зарастает с краев и превращается в болото, оно сменяется мокрым лугом, луг – кустарниками, а затем лесом. В зависимости от причин, вызвавших смену биоценоза, сукцессии делят на природные и антропогенные, аутогенные и аллогенные. Природные сукцессии происходят под действием естественных причин, не связанных с деятельностью человека. Антропогенные сукцессии обусловлены деятельностью человека. Аутогенные сукцессии (самопорождающиеся) возникают вследствие внутренних причин (изменения среды под действием сообщества); подтипы – сингенез и эндоэкокинез. Аллогенные сукцессии (порожденные извне) вызваны внешними причинами (например, изменение климата); подтипы – гейтогенез и гологенез. Сингенез – изменения происходят под влиянием взаимоотношений между видами сообщества, дифференциации экологических ниш без существенного изменения видами условий местообитания (первые стадии зарастания новых субстратов – песчаных отмелей, скальных осыпей, лавовых потоков и восстановление растительности на нарушенных человеком территориях – зарастание насыпей дорог, залежей и пр.). Эндоэкогенез – изменение сообщества путем изменения условий среды.Пример: процессы зарастания стариц, переход низинных болот в верховые и т.д. (природная сукцессия) и изменения под воздействием разного рода деструкций (полное или частичное разрушение сообщества в результате воздействия антропогенных факторов – рубки ухода за лесом). Гейтогенез – локальные (т.е. не связанные с общими тенденциями развития ландшафта) аллогенные смены, сравнительно быстрая по масштабу времени, вызываемая чаще всего антропогенными факторами (воздействие на растительность диких животных и хозяйственную деятельность человека – пастбищная дигрессия, воздействие мелиорации и пр.). Гологенез – изменения целых ландшафтов с более или менее параллельным изменением растительных сообществ и экосистем разных урочищ (развитие ильменей в низовьях Волги; строительства гидротехнических сооружений, вызывающих подтопление, затопление, засоление и пр.). В зависимости от первоначального состояния субстрата, на котором развивается сукцессия, различают первичные и вторичные сукцессии. Первичные сукцессии развиваются на субстрате, не занятом живыми организмами (на скалах, обрывах, сыпучих песках, в новых водоемах и т.п.). Вторичные сукцессии происходят на месте уже существующих биоценозов после их нарушения (в результате вырубки, пожара, вспашки, извержения вулкана и т.п.). В своем развитии экосистема стремится к устойчивому состоянию. Сукцессионные изменения происходят до тех пор, пока не сформируется стабильная экосистема, производящая максимальную биомассу на единицу энергетического потока. Сообщество, находящееся в равновесии с окружающей средой, называется климаксным. Принцип сукцессионного замещения – в процессе сукцессии формируется ряд экосистем, ведущий к наиболее устойчивой в данных условиях климаксовой экосистеме. Для развития биоценозов в ходе сукцессии характерен целый ряд общих закономерностей: постепенное увеличение видового разнообразия, смена доминирующих видов, усложнение цепей питания, увеличение в сообществах доли видов с длительными циклами развития, усиление взаимовыгодных связей в биоценозах и т. д.
Природные экосистемы В зависимости от природных и климатических условий можно выделить три группы и ряд типов природных экосистем (биомов). В основе классификации для наземных экосистем лежит тип естественной (исходной) растительности, для водных экосистем – гидрологические и физические особенности. Наземные экосистемы: Размещение по земной поверхности основных наземных биомов определяют два абиотических фактора – температура и количество осадков. Климат в разных районах земного шара неодинаков. Годовая сумма осадков меняется от 0 до 2500 мм и более. В связи с неравномерностью выпадения осадков различают влажные – гумидные (до 2000 мм/год) и засушливые (менее 250 мм/год) – аридные зоны; умеренные зоны расположены там, где выпадает промежуточное количество осадков (250–750 мм/год). При этом они выпадают равномерно в течение года или их основная доля приходится на определенный период – влажный сезон. Среднегодовая температура также варьирует от отрицательных величин до 38° С. Температуры могут быть практически постоянными в течение всего года (у экватора) или меняться по сезонам. 1. Тундры (в северном полушарии к северу от тайги). Климат очень холодный с полярным днем и полярной ночью, среднегодовая температура ниже 0° С. За несколько недель короткого лета земля оттаивает не более чем на один метр в глубину. Осадков менее 200-300 мм в год. Растительность: отсутствуют деревья, господствуют медленно растущие лишайники, мхи, травы (злаки и осоки), стелющиеся или карликовые кустарнички (брусника, черника) и кустарники (карликовая береза). Животный мир небогат, встречаются крупные травоядные копытные – северный олень (Евразия) и карибу (Северная Америка), мелкие роющие млекопитающие (лемминги), хищники (песец, горностай, ласка). Среди птиц преобладают полярная сова, ржанка, пуночка. Среди насекомых обильны двукрылые. Почвы тундровые – бедные с малой мощностью над слоем вечной мерзлоты. Очень ранимые экосистемы из-за медленного их восстановления. 2. Бореальные хвойные леса (тайга) (северные районы Евразии и Северной Америки). Климат: долгая и холодная зима, много осадков выпадает в виде снега. Растительность: господствуют вечнозеленые хвойные леса (ель, пихта, сибирская кедровая сосна, сосна обыкновенная, лиственницы) с мощной лесной подстилкой. Животный мир: крупные травоядные копытные (лось), мелкие растительноядные млекопитающие (барсук, белка, бурундук), хищники (медведь, рысь, росомаха, соболь, лисица, волк, норка), обилен гнус. Характерно множество болот и озер. Почвы подзолистые, дерново-подзолистые, мерзлотно-таежные – маломощные и бедные. 3. Листопадные леса умеренной зоны (широколиственные леса) (Западная Европа, Восточная Азия, восток США). Климат сезонный с зимними температурами ниже 0° С, осадков 750–1500 мм в год. Растительность: господствуют широколиственные листопадные породы деревьев (дуб, бук, клен, липа, ясень, граб), кустарниковый подлесок, мощная лесная подстилка. Животный мир: млекопитающие (лоси, медведи, рыси, лисицы, волки, белки, землеройки), птицы (дятлы, дрозды, совы, соколы). Биота адаптирована к сезонному климату: спячка, миграции, состояние покоя в зимние месяцы. Почвы бурые и серые лесные. В этих районах человеческая цивилизация получила наибольшее развитие, поэтому большая часть широколиственных лесов заменена культурными сообществами. 4. Степи умеренной зоны (в Евразии) и их аналоги: прерии (в Северной Америки), пампасы (в Южной Америки), туссоки (в Новой Зеландии). Климат сезонный, лето от умеренного теплого до жаркого, зимние температуры ниже 0° С, осадков 250–750 мм в год. Растительность: господствуют дерновинные злаки высотой до 2 м и выше в некоторых прериях Северной Америки или до 50 см, например, в степях России, с отдельными деревьями и кустарниками на влажных участках. Животный мир: крупные растительноядные млекопитающие – бизоны, вилорогие антилопы (Северная Америка), сайгаки, а ранее дикие лошади – тарпан (Евразия), кенгуру (Австралия), жирафы, зебры, белые носороги, антилопы (Африка); мелкие роющие млекопитающие (суслики, сурки, полевки, кролики), хищники (волки, койоты, львы, леопарды, гепарды, гиены), разнообразные птицы. Почвы: черноземы (самые плодородные почвы в мире) и каштановые. Большая часть степей в настоящее время используется под пашню, пастбища, сенокосы и т.д. 5. Чапараль (Средиземноморье, южный берег Австралии, в Калифорнии, Мексике и Грузии). Климат мягкий умеренный (средиземноморский – дождливая зима и засушливое лето), осадков 500-700 мм. Растительность: деревья и кустарники с жесткими вечнозелеными листьями (лавр, дуб, мирт, олеандр, можжевельник, дикая фисташка). Почвы коричневые и серо-коричневые. 6. Тропический грасленд и саванны (Центральная и Восточная Африка, Южная Америка, Австралия, значительная часть Южной Индии). Климат сухой и жаркий большую часть года, температура высокая круглый год, осадков 250–750 мм в год, распределяются неравномерно по сезонам (влажный и сухой сезоны). Растительность: травянистая растительность (злаковые) с редкими листопадными деревьями (баобабы, акации, пальмы). Животный мир: крупные растительноядные млекопитающие (антилопы, зебры, жирафы, носороги, слоны), хищники (львы, леопарды, гепарды), птицы (африканский страус, грифы). Много кровососущих насекомых, например, муха цеце. Почвы красные ферраллитные, красно-бурые и коричнево-красные. На распаханных землях выращивают злаковые, хлопчатник, арахис, сахарный тростник. 7. Пустыни травянистая и кустарниковая (некоторые районы Африки, например Сахара, Ближнего Востока и Центральной Азии, Большой Бассейн и юго-запад США, север Мексики и др.). Климат очень сухой, с жарким днем и холодными ночами, осадков менее 200–250 мм в год. Растительность: ксерофитные травы и редкостойный кустарник, кактусы, множество эфемеров, быстро развивающихся после непродолжительных дождей. Корневые системы у растений обширные, поверхностные, перехватывающие влагу редких осадков или стержневые корни, проникающие в землю до уровня грунтовых вод (30 м и глубже). Животный мир: разнообразные грызуны (тушканчики, суслики), копытные (куланы, джейран, вилорогая антилопа), хищники (волк, койот, корсак). Из птиц саджа, рябки, жаворонки. Много пресмыкающиеся, насекомых и паукообразных. Почвы светло-бурые, сероземы. Экосистемы хрупкие, легко нарушаются в результате перевыпаса, ветровой и водной эрозии. 8. Полувечнозеленые сезонные листопадные тропические леса (тропическая часть Азии, Центральная Америка). Климат со сменой сухого (4–6 месяцев) и влажного сезонов, среднегодовое количество осадков 800–1300 мм в год. Растительность: господствуют леса. Доминируют деревья верхнего яруса, сбрасывающие листья в сухой сезон. Нижний ярус образуют в основном вечнозеленые деревья и кустарники. Много лиан и эпифитов. Животный мир: практически так же богат, как в вечнозеленых тропических дождевых лесах. Характерны слоны, жирафы, буйволы. Почвы красные ферраллитные. 9. Вечнозеленые тропические дождевые леса (север Южной Америки, Центральная Америка, западная и центральная части экваториальной Африки, Юго-Восточная Азия, прибрежные районы северо-запада Австралии, острова Индийского и Тихого океанов). Климат без смены сезонов в связи с близостью к экватору, среднегодовая температура выше 17°С (обычно 28° С), среднегодовое количество осадков превышает 2000–2500 мм в год. Растительность. Деревья разной высоты образуют густой полог из многих ярусов (выделяют до 10–12 ярусов). Кустарники отсутствуют, травяной покров беден. На стволах и ветвях деревьев развиваются растения-эпифиты, корни которых не достигают почвы, и деревянистые лианы, укореняющиеся в почве и взбирающиеся по деревьям до их вершин. Видовое разнообразие растений огромно. Животный мир по видовому составу богаче, чем во всех других биомах вместе взятых: млекопитающие (обезьяны, ленивцы, ягуары), птицы (попугаи, колибри, туканы). Встречаются многочисленные пресмыкающиеся, земноводные, насекомые с яркой окраской. Почвы красно-желтые ферраллитные – маломощные и бедные органическим веществом и минеральными элементами питания растений. Большая часть питательных веществ закреплена в биомассе растительности. При сведении тропических лесов под пашню почвы теряют плодородие за 2–3 года. Пресноводные экосистемы: Распределение организмов в водных экосистемах зависит от степени освещенности. Выделяются следующие зоны: литоральная зона (толща воды, где солнечный свет доходит до дна), лимническая зона (толща воды до глубины, куда проникает всего 1% от солнечного света и где затухает фотосинтез), эвфотическая зона (вся освещенная толща воды – включает литоральную и лимническую зоны), профундалъная зона (дно и, толща воды, куда не проникает солнечный свет). В проточных водоемах выделяют перекаты (мелководные участки с быстрым течением: дно без ила, встречаются преимущественно прикрепленные формы перифитона и бентоса) и плесы (глубоководные участки: течение медленное, на дне мягкий илистый субстрат и роющие животные). По количеству проникающего света водоемы подразделяют на две горизонтальные зоны: верхнюю или эфотичесую (до 100–200 м) и нижнюю, простирающуюся до больших глубин – афотическую, где света для фотосинтеза недостаточно. 1. Лентические экосистемы (озера, пруды, водохранилища и др.). Литоральная зона населена двумя группами растений: укрепившиеся в дне (камыши, рогозы, кувшинки, прикрепленные водоросли и др.) и плавающие (водоросли, рдесты и др.). Животные в литорали более разнообразны, чем в других зонах водоема. Встречаются моллюски, коловратки, мшанки, личинки насекомых и др. Рыбы большую часть жизни проводят в литорали и здесь же размножаются. Многие обитающие здесь животные дышат кислородом атмосферного воздуха (лягушки, саламандры, черепахи и др.). Зоопланктон представлен ракообразными, имеющими большое значение для питания рыб (дафнии и др.). Лимническая зона. Продуценты представлены фитопланктоном. В водоемах умеренного пояса «цветение» весной связано с массовым развитием диатомовых, летом – зеленых, осенью – азотфиксирующих сине-зеленых водорослей. Зоопланктон представлен растительноядными ракообразными и коловратками. Нектон лимнической зоны – только рыбы. Профундальная зона около дна представлена бентосными формами – личинками насекомых, моллюсками, кольчатыми червями, сапротрофными бактериями и грибами. 2. Лотические экосистемы (реки, родники, ручьи и др.) отличаются от стоячих водоемов следующими особенностями: 1) наличие течения; 2) более активный обмен между водой и сушей; 3) более высокое содержание кислорода и более равномерное его распределение; 4) преобладание детритных цепей питания (здесь более 60% энергии консументы получают от привнесенного материала). Выделяют логические сообщества перекатов и плесов. На перекатах поселяются организмы, способные прикрепиться к субстрату (например, нитчатые водоросли) или хорошие пловцы (например, форель). На участках плеса сообщества напоминают прудовые. В больших реках прослеживается продольная зональность: в верховьях – сообщества перекатов, в низовьях и дельте – плесов, между ними местами могут возникать и те и другие. Видовой состав рыб к низовьям обедняется, но увеличиваются их размеры. 3. Заболоченные участки и болота бывают низинные (имеют, как правило, питание подземными водами) и верховые (питаются атмосферными осадками). Верховые могут встречаться в любом понижении или даже на склонах гор, низинные возникают вследствие зарастания озер и речных стариц. Здесь распространены болотные растения. Болотные почвы и торфяники содержат много углерода. Их сельскохозяйственная отработка приводит к выделению в атмосферу большого количества углекислого газа. Морские экосистемы: 1. Открытый океан беден биогенными элементами. Эти районы можно считать «пустынями» по сравнению с прибрежными водами. Арктические и антарктические зоны более продуктивны, так как плотность планктона растет при переходе от теплых морей к холодным, и фауна рыб и китообразных здесь значительно богаче. Продуцентом выступает фитопланктон, им питается зоопланктон, а тем в свою очередь нектон. Видовое разнообразие фауны снижается с глубиной. На глубине в стабильных местообитаниях сохранились виды из далеких геологических эпох. 2. Глубоководные рифтовые зоны океана находятся на глубинах около 3000 м и более. Условия жизни в экосистемах глубоководных рифтовых зон очень своеобразны. Это полная темнота, огромное давление, пониженная температура воды, недостаток пищевых ресурсов, высокая концентрация сероводорода и ядовитых металлов, встречаются выходы горячих подземных вод, и т. д. В результате живущие здесь организмы претерпели следующие адаптации: редукция плавательного пузыря у рыб или заполнение его полости жировой тканью, атрофирование органов зрения, развитие органов светосвечения и др. Живые организмы представлены гигантскими червями (погонофорами), крупными двустворчатыми моллюсками, креветками, крабами и отдельными видами рыб. Продуцентами выступают сероводородные бактерии, живущие в симбиозе с моллюсками. 3. Область континентального шельфа является самой богатой в фаунистическом отношении. Прибрежная зона очень благоприятна по условиям питания, даже в дождевых тропических лесах нет такого разнообразия жизни, как здесь. 4. Районы апвеллинга расположены вдоль западных пустынных берегов континентов. Здесь наблюдается апвеллинг – подъем холодных вод с глубины океана, так как ветры перемещают воду от крутого материкового склона, а взамен ей из глубины поднимается вода, обогащенная биогенными элементами. Эти районы богаты рыбой и птицами, живущими на островах. 5. Эстуарии, лиманы, устья рек, прибрежные бухты и т. д. – прибрежные водоемы, представляющие собой экотоны между пресноводными и морскими экосистемами. Это высокопродуктивные районы, где наблюдается аутвеллинг – привнесение биогенных элементов с суши. Они обычно входят в литоральную зону и подвержены приливам и отливам. Здесь встречаются болотные и морские травы, водоросли, рыба, крабы, креветки, устрицы и т.д. 6.9. Антропогенные экосистемы Агроэкосистемы(сельскохозяйственные экосистемы, агроценозы) – искусственные экосистемы, возникающие в результате сельскохозяйственной деятельности человека (пашни, сенокосы, пастбища). Агроэкосистемы создаются человеком для получения высокой чистой продукции автотрофов (урожая). В них, так же, как в естественных сообществах, имеются продуценты (культурные растения и сорняки), консументы (насекомые, птицы, мыши и т.д.) и редуценты (грибы и бактерии). Обязательным звеном пищевых цепей в агроэкосистемах является человек. Отличия агроценозов от естественных биоценозов: – незначительное видовое разнообразие (агроценоз состоит из небольшого числа видов, имеющих высокую численность); – короткие цепи питания; – неполный круговорот веществ (часть питательных элементов выносится с урожаем); – источником энергии является не только Солнце, но и деятельность человека (мелиорация, орошение, применение удобрений); – искусственный отбор (действие естественного отбора ослаблено, отбор осуществляет человек); – отсутствие саморегуляции (регуляцию осуществляет человек) и др. Таким образом, агроценозы являются неустойчивыми системами и способны существовать только при поддержке человека. Сельскохозяйственное освоение территорий часто приводит к разрушению созданных природных механизмов регуляции численности отдельных видов и резким изменениям в уровне их обилия. В борьбе человека с сорняками и вредителями культурных растений постоянно возникает экологический эффект бумеранга . В современном сельском хозяйстве в изобилии применяют разнообразные химические средства защиты растений и животных – пестициды (от лат. pestis – зараза и caedo – убиваю) (ядохимикаты). В группу пестицидов включают: – гербициды (от лат. herba – трава и caedo – убиваю) – химические препараты для уничтожения нежелательной, главным образом сорной растительности (гербициды сплошного действия поражают все виды растений, используются вокруг промышленных объектов, на аэродромах, под линиями электропередач и др.; избирательного – уничтожают одни виды, например, сорняки, и не повреждают культурные растения, применяют в сельском хозяйстве для химической прополки на полях, в садах, виноградниках и т. п.); – инсектициды (от лат. insectum – насекомое и caedo – убиваю) – химические препараты для борьбы с насекомыми – вредителями сельскохозяйственных растений; – акарициды (от греч. akari – клещ и лат. caedo – убиваю) – химические препараты из группы пестицидов для уничтожения клещей, вредных для сельскохозяйственных растений и животных; – зооциды (от греч. zoon – животное и лат. caedo – убиваю) – химические препараты, использующиеся главным образом для борьбы с грызунами (родентициды, включают ратициды для уничтожения крыс); фунгициды (от лат. fungus – гриб и caedo – убиваю) – химические препараты для уничтожения или предупреждения развития патогенных грибов – возбудителей болезней сельскохозяйственных растений; – дефолианты – химические препараты, вызывающие искусственный листопад, что ускоряет созревание и облегчает уборку урожая (применяют для предуборочного опадения листьев, в основном у хлопчатника); – десиканты – химические препараты, вызывающие обезвоживание тканей растений, что ускоряет их созревание и облегчает уборку урожая (применяются для подсушивания хлопчатника, риса, клещевины, картофеля и др.); – регуляторы роста растений. Большинство пестицидов не обладает избирательным действием и подавляет не только те виды, против которых применяется, но и их паразитов и хищников. Таким образом, нарушаются возникающие в агроценозах регуляторные связи. Хищники и паразиты, занимая более высокие уровни в цепях питания, более чувствительны к ядам, чем те виды, которыми они питаются. Оставшаяся часть вредителей, освобожденная от регуляторов, дает новую, еще более высокую вспышку численности. Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 706; Нарушение авторского права страницы
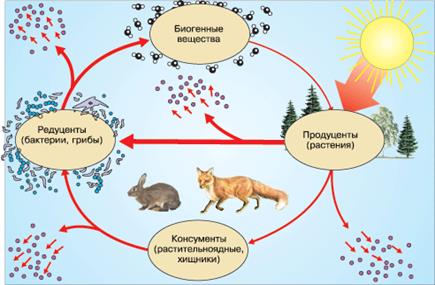 Таким образом, как правило, в любой экосистеме можно выделить три функциональные группы организмов: продуцентов, консументов и редуцентов. В экосистемах, образованных только микроорганизмами, консументы отсутствуют. В каждую группу входит множество популяций разных видов, населяющих экосистему. В экосистеме пищевые и энергетические связи идут в направлении: продуценты → консументы → редуценты.
Таким образом, как правило, в любой экосистеме можно выделить три функциональные группы организмов: продуцентов, консументов и редуцентов. В экосистемах, образованных только микроорганизмами, консументы отсутствуют. В каждую группу входит множество популяций разных видов, населяющих экосистему. В экосистеме пищевые и энергетические связи идут в направлении: продуценты → консументы → редуценты.