Вопрос 4Течения Индийского океана
Физические характеристики течения
- Скорость течения колеблется от 24 до 80 миль в сутки. Средний показатель скорости около 40 миль в сутки.
- Температура воды — н/д
- Солёность — н/д
Вопрос 1 Плотность вод мирового океана и особенности ее пространственного распределения
Плотность океанических вод повсеместно увеличивается от поверхности ко дну. При этом вначале, примерно до глубины 1000 - 1500 м, плотность повышается быстро, а затем медленно, едва заметно.
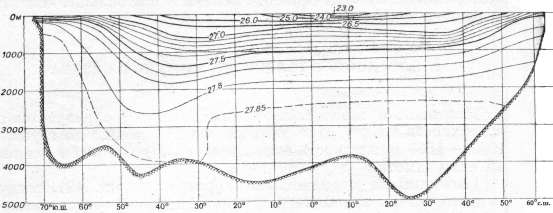
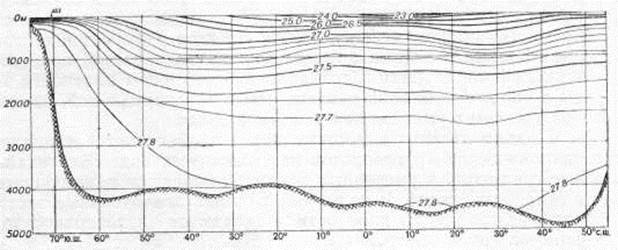
Изопикны на меридиональных разрезах, следуя за изотермами, приподняты в экваториальной зоне в связи с восходящими потоками, приносящими к поверхности более плотные глубинные воды. В результате этого здесь создаются наибольшие в Мировом океане вертикальные градиенты плотности. Преобладание нисходящих потоков в низких широтах, вызывающееся антициклоническими круговоротами, приводит к опусканию изопикн. Вертикальные градиенты плотности воды в тропической зоне меньше, чем в экваториальной, однако больше, чем в умеренной и полярной зонах.
Условная плотность воды по меридиональному сечению Атлантического океана (по средним широтным величинам)
Условная плотность воды по меридиональному сечению Тихого океана (по средним широтным величинам)
Пространственное распределение океанографических
Характеристик
К основным океанографическим характеристикам, обуславливающим поведение и воспроизводство объектов рыболовного промысла, относятся: температура, соленость и плотность вод Мирового океана, а также перемещение водных масс в трехмерной системе течений. Волновые процессы, прозрачность воды, содержание кислорода могут быть отнесены к факторам, влияющим на перемещение рыбных скоплений, но комплекс первых названных свойств образует среду с основными физическими и динамическими предпосылками, формирующими остальные условия обитания живых организмов в море.
Как известно, приток тепла к определенной точке морской поверхности обусловлен, главным образом, углом падения солнечных лучей относительно линии от данной точки горизонта. Количество тепловой энергии, проникающее в воду, зависит от прозрачности ее, следовательно, уменьшается с глубиной. По Свердрупу доля суммарной солнечной энергии снижается в процентном отношении от исходной следующим образом:
| Глубина (м)
|
|
|
|
|
|
|
|
| | Суммарная солнечная радиация %
|
| 37, 7
| 31, 6
| 23, 7
| 16, 1
| 9, 35
| 2, 69
| 0, 45
|
В прибрежных водах при увеличении мутности воды ослабление солнечной радиации происходит еще быстрее:
| Глубина (м)
|
|
|
|
|
|
|
| | Суммарная солнечная радиация %
|
| 26, 7
| 17, 0
| 5, 95
| 1, 21
| 0, 05
| ---
|
При распространении тепла в глубинные слои только под воздействием молекулярной диффузии, определенное значение температуры от поверхности до 100 м передалось бы только через 2000 лет. Однако, турбулентное перемешивание, обусловленное ветровым волнением и анизотропностью морских течений, увеличивает скорость передачи тепла по вертикали на несколько порядков. Конвективное перемешивание вод при выхолаживании в зимний период в умеренных и полярных широтах обеспечивает выравнивание температуры верхнего слоя океана за счет переноса холода в нижние слои. Плотностная конвекция при испарении и осолонении поверхностных вод наоборот переносит в глубину тепло.
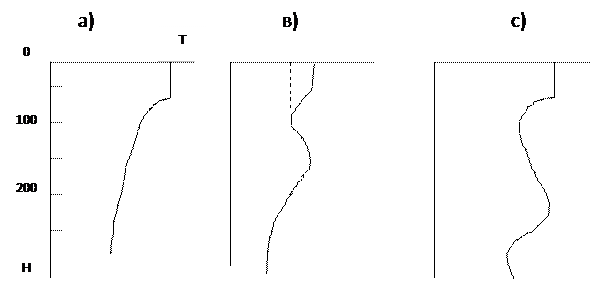 В результате вышеописанных процессов формируется профиль вертикального распределения температуры (Рис. 1) В результате вышеописанных процессов формируется профиль вертикального распределения температуры (Рис. 1)
Рис. 1. Вертикальные профили температуры воды в море в разных широтных зонах: а - тропической, в – умеренной, с – полярной.
Как видим – общее генеральное изменение температуры отражает ее характерное падение с глубиной. На эту закономерность накладывается ряд особенностей, обусловленных широтными климатическими условиями и адвекцией вод сопредельных районов. В тропических широтах (Рис. 1, а), где отсутствует плотностная конвекция при зимнем выхолаживании, весь год сохраняется верхний квазиоднородный слой толщиной 10-50 м, образованный постоянным прогревом и ветровым перемешиванием. Ниже находится слой повышенного вертикального градиента температуры, именуемый слоем скачка или термоклином. В умеренных широтах (Рис. 1, в) такой же термоклин и верхний квазиоднородный слой образуются в летний период. При осенне-зимнем выхолаживании эта структура разрушается и температура выравнивается по вертикали в слое от 100 до 400 м в результате плотностной конвекции. Последующий летний прогрев и ветровое перемешивание в некоторых морях (Черное, Охотское) не ликвидируют подповерхностную холодную, так называемую «зимнюю», воду и тем самым сохраняется промежуточный минимум температуры. В полярных широтах (Северный Ледовитый океан, воды Арктики) образуется промежуточный максимум температуры (Рис. 1, с), который образуется проникновением более соленых и теплых вод субтропической зоны в полярные районы. Характерным примером является глубинный слой относительно теплых вод, простирающийся от зоны субтропической конвергенции почти до шельфа Антарктиды.
Изменение температуры поверхностного слоя океана в основном соответствует широтным зонам, аналогично распределению солнечной энергии на поверхности земного шара. В южном полушарии, где сезонный нагрев и выхолаживание материков в меньшей степени искажают характерное распределение температуры по сравнению с северным полушарием, изотермы проходят почти параллельно широтам. В северном полушарии, кроме того, система разгрузки теплых вод, избыток которых образуется у восточных берегов материков в тропических широтах, возникающий из-за «пассатного нагона», создает меридиональные потоки в виде теплых течений, например Гольфстрим в Атлантическом океане и Куросио в Тихом. Приток вод в полярные широты компенсируется встречными холодными течениями – соответственно Лабрадорским и Ойясио. Таким образом, распределение температуры в северном полушарии получается более сложным и с большими различиями между холодными и теплыми половинами года.
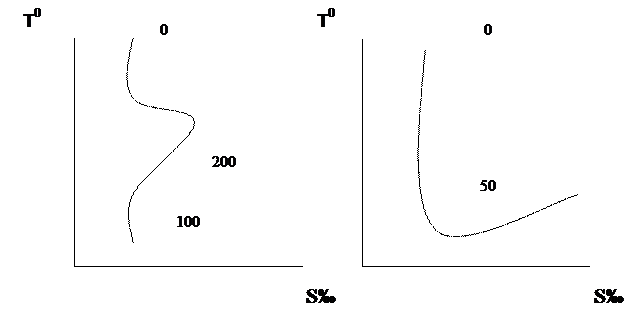
Температура вместе с соленостью воды определяет ее плотность. Зависимость плотности от температуры обратная (чем холоднее вода, тем она плотнее), а от солености – прямая. Поскольку вертикальная стратификация водных масс обусловлена увеличением плотности с глубиной, то уменьшение температуры и увеличение солености от поверхности до дна является общей закономерностью. Характерный вертикальный профиль роста солености с глубиной искажается рядом причин. Интенсивное испарение вод в тропических и субтропических широтах создает запас соленых вод, которые погружаются на нижележащие горизонты и создают там промежуточные максимумы. Наиболее интенсивными «испарителями» являются Средиземное и Красное моря, где соленость превышает 40%. Погруженные средиземноморские воды в Атлантическом океане и красноморские в Индийском, создают глубинные слои повышенной солености (Рис. 2). В то же время, перемешивание соленых вод субтропических широт с относительно распресненными и холодными водами полярных создает подповерхностные слои промежуточного минимума солености.
| | | |  |
Рис. 2. Вертикальный профиль солености в субтропических широтах.
В районах стока пресных вод в море в верхнем слое создается обычно повышенный вертикальный градиент солености, именуемый халлоклином. В Черном море, например, халлоклин существует на глубинах 75-120 м и обуславливает здесь мощный градиент плотности (пикноклин), который в отличие от сезонного термоклина, не разрушается зимней конвекцией. Это обстоятельство создает препятствие аэрации нижележащих вод и является причиной образования сероводородной зоны от 100-150 метров до дна.
Пространственное распределение солености определяется водным балансом осадков и испарения плюс влияние ледотаяния. Максимальная соленость в океане наблюдается в тропических и субтропических широтах – до 36%. Наименьшая соленость 30-34% характерна для высоких широт. В экваториальной зоне также существует пояс пониженной солености (33-34‰), которая создается интенсивными осадками. Естественно, что в районах впадения рек в океан возрастание солености начинается с нулевого значения.
Плотность морской воды (в г/см3) отличается от плотности пресной на 0, 02-0, 03 грамма в кубическом сантиметре. Для расчетов и построения графиков используется условная плотность: Gt=(r-1)*103, таким образом, если r=1, 01425, то st=14, 25.
Распределение плотности по вертикали определяется условием стратификации, при которой более тяжелая вода не может находиться выше легкой. Увеличение этой характеристики с глубиной также не является равномерным. Конвекция при зимнем выхолаживании или интенсивном испарении и осолонении, а также ветровое перемешивание, создают слой повышенного вертикального градиента плотности (пикноклин), совпадающий с термо- или халлоклином, или сочетанием обеих характеристик. В открытом океане сезонный пикноклин располагается в пределах от 25 до 100 м, его глубина в общем увеличивается от полярных районов к тропическим. В соответствии с распределением температуры и солености в разных районах океана формируются воды различной плотности: в низких широтах менее плотные, в высоких – более плотные. Это обусловлено преобладающим влиянием температуры.
 Сочетание определенных значений температуры и солености является основной характеристикой водных масс. Водные массы представляют собой большие, сравнимые с размерами океана, объемы вод, формирующиеся в квазистационарных климатических условиях. Каждая из них в конкретной географической точке может быть представлена отрезками кривой, проведенной в координатах температуры и солености. Это так называемые Т, S – кривые, принятые в океанографии для изображения распределения водных масс в любой точке моря с наблюдениями за температурой и соленостью (Рис.3). На базе множества точек в Т, S – поле строятся карты водных масс, а с применением объемного статистического анализа определяются их объемы, количества в них тепла и солей. Сочетание определенных значений температуры и солености является основной характеристикой водных масс. Водные массы представляют собой большие, сравнимые с размерами океана, объемы вод, формирующиеся в квазистационарных климатических условиях. Каждая из них в конкретной географической точке может быть представлена отрезками кривой, проведенной в координатах температуры и солености. Это так называемые Т, S – кривые, принятые в океанографии для изображения распределения водных масс в любой точке моря с наблюдениями за температурой и соленостью (Рис.3). На базе множества точек в Т, S – поле строятся карты водных масс, а с применением объемного статистического анализа определяются их объемы, количества в них тепла и солей.
Рис. 3. Т, S – кривые: а – тропические широты, в – Черное море.
Плотность морской воды в среднем уменьшается от высоких широт (27, 5 у.е.) к экватору, где достигает самых низких, для открытого океана, величин (21, 0 – 22, 0 у.е.). Средняя условная плотность Мирового океана 24, 74, самая низкая (18, 0) отмечена в Панамском заливе (Тихий океан), самая высокая (28, 0) – юго-западнее Шпицбергена и в водах Антарктики. В полярных морях плотность воды относительно высокая (в Норвежском, Гренландском, Баренцевом) из-за низкой температуры и проникновения соленых вод Атлантики. В Красном море и Персидском заливе она высока из-за большой солености. В Балтийском море, в Азовском плотность низкая по причине речного стока. В Черном море изменение солености по глубине существенное – от 18‰ на поверхности до 22‰ ниже 100 м, поэтому галлоклин на этих глубинах образует мощный пикноклин, который не разрушается зимней температурной конвекцией.
| Вопрос 2 Факторы определяющие распределение живых организмов в водах Мирового океана.
| | Организм постоянно подвергается действию изменчивых факторов окружающей среды. Было уже отмечено, что это приводит к формированию экологической валентности вида, которая является отражением степени действия экологических факторов. Следовательно, анализ сообществ организмов способен давать объективную информацию о состоянии окружающей среды. Мы рассмотрим способность организмов отражать параметры разных сред.
1. Общие положения. Известно, чем сильнее изменяется данный параметр среды в пространстве или во времени, тем обычно большее экологическое значение он имеет для организмов. Существенную роль при этом играют и степень регулярности воздействия данного фактора на организмы, регулярность изменяемости фактора во времени. В соответствии с этим среди изменчивых факторов можно различать те, которые изменяются закономерно периодически и без закономерной периодичности. Факторы, изменяющиеся периодически, обусловливают формирование у видов характерных для них биологических циклов (суточных, сезонных, годовых и т.д.), специфических типов динамики численности некоторых других видов признаков. Факторы, изменяющиеся без закономерной периодичности, главным образом влияют на распространение вида и его численность. Факторы, имеющие периодический и регулярный характер, можно разделить на первичные и вторичные. К первым относятся воздействия, являющиеся прямым следствием ритмики геофизических циклов. Вторичные – это воздействия, являющиеся косвенным следствием ритмики геофизических циклов (например, паводковые и межпаводковые периоды).
Особи каждого вида характеризуются определенным типом обмена веществ и энергии, без сохранения которого не могут успешно развиваться. Если состояние среды таково, что организму грозит нарушение баланса обмена веществ и энергии, то он либо изменяет положение в пространстве, приводя себя по возможности в оптимальные условия среды, либо меняет интенсивность обмена или даже характер последнего.
2. Оптимизация экологических условий. Оптимальными называются условия, в которых организм с наименьшими энергетическими затратами сохраняет характерный для него тип обмена веществ. Оптимальные значения какого-либо фактора применительно к различным функциям организма различны, поэтому оптимум в отношении организма в целом – понятие интегральное. Оптимальные условия – это не те, которые обеспечивают наилучшие условия для протекания отдельных функций, а те, при которых суммарный эффект протекания всех функций является наилучшим с точки зрения обеспечения биологического процветания вида. Оптимальная дозировка фактора для каждого организма представляет собой величину не постоянную, а переменную, зависящую от состояния самого организма и от всей суммы условий, в которых он обитает.
Не все факторы водной среды играют равную роль в жизни организмов. В связи с этим они делятся на первостепенные и второстепенные. К важнейшим из них относятся механические и динамические свойства воды и донного грунта, температура, свет, растворенные и взвешенные в воде вещества, активная реакция среды и некоторые другие.
3. Факторы водной среды. Общая характеристика. Водная среда жизни, гидросфера, занимающая до 71% площади земного шара, включает около 1, 46 млрд км3 воды. Основной объем воды (примерно 95%) сосредоточен в Мировом океане, львиная доля пресных вод – в ледниках (85%) и подземных водах суши (14%), на озера, водохранилища, пруды, болота, реки и ручьи приходится чуть более 0, 6% от общего объема пресной воды, остающиеся 0, 35% заключены в почвенной влаге и парах атмосферы.
В водной среде обитают около 150 000 видов животных (примерно около 7% от общего количества на Земле) и 10 000 видов растений (8%). Следовательно, вода как среда жизни не отличается видовым разнообразием.
В морях и океанах животный и растительный мир наиболее разнообразен и богат в экваториальной и тропической зонах. С удалением от этих поясов на север и юг качественный состав организмов постепенно беднеет. Основная масса организмов Мирового океана сосредоточена у берегов, преимущественно в зоне морских побережий. Открытые воды, расположенные вдали от берегов, представляют собой пустынные области, практически лишенные жизни.
Доля рек, озер, болот по сравнению с морями и океанами в биосфере незначительна. Несмотря на меньшее биоразнообразие в пресных водах и их небольшой объем, они имеют выдающееся значение для огромного количества животных и растений, а также для человека, обеспечивая их необходимым запасом пресной воды.
Современную гидросферу можно рассматривать не только как среду жизни. Ее обитатели оказывают активное влияние на круговорот веществ в биосфере.
Свойства воды. Значение воды в жизнедеятельности организмов определяется главным образом ее физическими свойствами. Среди этих свойств, прежде всего, выделяются термические – большая теплоемкость, высокая скрытая теплота плавления и испарения, низкая теплопроводность, расширение перед замерзанием. Благодаря этим свойствам поддерживается относительное постоянство температурного режима океанов, что в свою очередь, уменьшает амплитуду колебаний температуры на земной поверхности. Расширение воды перед замерзанием и аномальные изменения плотности воды в интервале от 0 до +4°С (именно при такой температуре вода имеет наибольшую плотность, при дальнейшем повышении или понижении температуры воды ее плотность уменьшается) обеспечивают перемешивание водных масс и препятствуют промерзанию водоемов. Не будь этих аномалий образующийся в холодное время года лед опускался бы на дно водоемов, превращая водные бассейны в залежи льда, оттаивающие летом лишь с поверхности, где находили бы приют только эфемерные водные организмы.
Вода является превосходным растворителем. Это свойство воды и ее исключительная подвижность делают воду основным фактором обмена веществ, как в неорганической, так и органической природе. В растворенном состоянии органические и неорганические вещества поступают к потребителям. Водой транспортируются вещества внутри организмов, с водой выделяются продукты распада.
Благодаря высокому поверхностному натяжению воды (по ее поверхности способны бегать водомерки), она удерживается на поверхности живых и неживых объектов и поднимается по капиллярам. Именно это свойство воды обеспечивает питание наземных растений.
Практическая несжимаемость воды позволяет организмам населять большие глубины.
Благодаря ряду оптических свойств прежде всего, прозрачности, в воде на значительных глубинах может идти фотосинтез.
Механико-динамические свойства воды и грунта определяют условия фиксации и передвижения гидробионтов в пространстве. Они обуславливают условия плавания пелагических организмов и их возможности удержания в тех или иных пространственных участках водоема. Бентосные организмы в зависимости от механико-динамических свойств грунта обладают разными возможностями закапывания в субстрат, фиксации на нем и передвижения. Недостаточная стабильность грунта – препятствие для существования бентосных организмов (невозможность строительства убежищ, механические повреждения, вымывания в толщу воды, захоронение под толщей осаждающихся осадков и т.д.). Движение воды – вынос за пределы биотопов с благоприятными условиями обитания и наоборот.
Экологические группы водных организмов. На основе образа жизни среди водных организмов выделяют несколько групп:
– бентос (от греч. benthos – глубина) – прикрепленные к грунту, лежащие на нем или живущие в толще осадков организмы. Представлен фитобентосом, зообентосом и бактериобентосом. В зообентосе по способу питания выделяют хищников, фильтраторов, грунтоедов и т.д.;
– перифитон (от греч. peri – вокруг, возле) – животные и растения, либо прикрепленные, либо удерживающиеся за стебли и листья высших растений или любые поверхности, возвышающиеся над дном и плавающие по течению;
– планктон (от греч. planktos – парящий, блуждающий и on – сущее) – пассивно плавающие растительные (фитопланктон) или животные (зоопланктон) организмы, перемещающиеся главным образом благодаря течениям;
– нектон (от греч. nektos – плавающий) – активные пловцы с обтекаемой формой тела и развитыми органами движения, не связанные непосредственно с дном (рыбы, кальмары, ластоногие, киты и т.д.);
– нейстон (от греч. neustos – плавающий) – сообщество микроорганизмов, растений и животных мелких и средних размеров, обитающих у поверхности воды на границе водной и воздушных сред (над или под поверхностной пленкой, до 5 см в глубь воды). Это бактерии, простейшие, клопы-водомерки, водоросли, личинки некоторых организмов;
– плейстон (от греч. pleystikos – плавающий) – совокупность гидробионтов, часть тела которых находится в воде, а часть – над ее поверхностью. К ним относятся парусники, некоторые сифонофоры и членистоногие, ряска.
Обитатели рек носят название потамобионтов. Те из них, что живут только на течении, называются реофилами. Движение воды в реках вызывает эрозию ее ложа, идущую в глубинном и боковом направлениях. Перенос и седиментация грунта изменяют конфигурацию ложа в горизонтальном и вертикальном направлениях. В результате боковой эрозии река меандрирует, образуя излучины, или меандры. Когда река спрямляет свое русло, тогда отшнуровавшиеся излучины превращаются в старицы. В других случаях от реки могут обособляться, не теряя с нею связи, закосья, затоны и протоки.
По биологической классификации, предложенной А. Тинеманном и Е. Науманном, различают озера эвтрофные, олиготрофные и дистрофные (eu – хорошо, oligos –мало, dis – недостаточно, trophos – пища). К эвтрофным (высококормным) относятся неглубокие равнинные озера с хорошо выраженной литоралью и богатой растительностью. Их вода богата питательными солями, грунты содержат много органического вещества, и на них развивается богатая донная фауна. Олиготрофные (малокормные) озера обычно бывают рсположены на кристаллических породах, обладают значительной глубиной и слаборазвитой литоралью. Донные отложения бедны органическими веществами, в воде содержится мало питательных солей, соответственно, жизнь на дне и в толще воды отличается бедностью. Дистрофные (недостаточно кормные) озера представляют собой неглубокие заболоченные водоемы с торфянистыми отложениями на дне. Торфянистые отложения исключают контакт воды с грунтом и поэтому в ней содержится очень мало минеральных веществ, солей, нужных для питания растений. Планктон и бентос в этих озерах развит очень слабо. Обитатели озер называются лимнобионтами. В соответствии с членением бентали выделяются формы.
Активная реакция среды представляет собой свойство, обусловливающееся присутствием ионов Н+ и ОН-. Как известно, часть молекул воды диссоциирует на эти ионы. Если концентрации ионов Н+ и ОН- равны, вода будет нейтральной. С увеличением содержания ионов Н+ и ОН- вода будет соответственно кислой (Н+) или щелочной (ОН-). Обычно в качестве показателя активной реакции берется не концентрация Н+, а ее десятичный логарифм с обратным знаком. Эта величина, называемая водородным показателем, обозначается символом рН. Если рН меньше 7, вода кислая, больше 7 – щелочная, для нейтральной воды рН равен 7. Степень диссоциации воды зависит от температуры: с понижением последней содержание Н+ падает, а с повышением соответственно увеличивается. Следовательно, заключение об активной реакции воды надо делать с учетом температуры.
Активная реакция природных вод довольно устойчива. Тем не менее в одном и том же водоеме рН в течение суток может колебаться на 2 единицы и более: ночью понижаться в результате подкисления воды выделяющимся в процессе дыхания углекислым газом, днем повышается за счет потребления СОя фотосинтезирующими растениями. В грунтах озер и болот рН обычно несколько ниже 7, в океанических осадках он часто бывает сдвинут в щелочную сторону. В зависимости от отношения к различным концентрациям водородных и гидроксильных ионов гидробионты подразделяются на стеноионных, обитающих в водах с колебанием рН в 5–6 единиц, и эвриионных, выдерживающих большие изменения этого фактора.
Концентрация водородных ионов не только определяет границы распространения гидробионтов, но и влияет на характер их жизнедеятельности.
Окислительно-восстановительный потенциал характеризует собой условия протекания в среде окислительных и восстановительных процессов, от которых зависят многие стороны жизнедеятельности гидробионтов особенно микроорганизмов. Напомню, что окислительно-восстановительная реакция заключается в том, что одно вещество, отдавая свои электроны и заряжаясь положительно, окисляется, а другое, приобретая электроны и заряжаясь отрицательно, восстанавливается. В результате между ними возникает разность электрических потенциалов. Чем больше эта разность, тем выше окислительная способность среды. Она определяется концентрацией молекулярного водорода в воде, а также состоянием таких равновесных систем, как Fe (3) – Fe (2), Mn (4) – Mn (2), S (2) – S(1), а также другими элементами с переменной валентностью.
Таким образом, условия жизни в воде своеобразны. Большое влияние на распределение организмов оказывают свет, температура, течения, давления, активная реакция среды, растворенные газы (в первую очередь кислород) и соли. Морские и континентальные воды резко отличаются по условиям жизни. Морская вода представляет собой более благоприятную среду, близкую к физиологически уравновешенному раствору. Поэтому ее обитатели осмотически открыты. Континентальные воды обладают неблагоприятными условиями жизни, их обитатели осмотически закрыты.
Адаптивные особенности водных растений. Водные растения в отличие от наземных поглощают влагу и минеральные соли непосредственно из окружающей воды, поэтому их организация имеет свои особенности. У них слабо развиты проводящие ткани, а также корневая система. Поскольку корни служат в основном для прикрепления к подводному субстрату, они лишены корневых волосков. Мощное развитие корневой системы у некоторых из них – кувшинок, кубышек – обеспечивает вегетативное размножение и запасание некоторых веществ.
Главной структурной особенностью гидрофитов является наличие крупных межклетников и полостей, создающих особую воздушную ткань, которая обеспечивает плавучесть органов. Подводные гидрофиты отличаются от надводных отсутствием функционирующих устьиц, тонкими рассеченными листьями, слабым развитием механических тканей. Интенсивный газообмен при недостатке в воде растворенного кислорода обеспечивается либо очень длинными и тонкими стеблями и листьями, покровы которых легко проницаемы для кислорода, либо сильной расчлененностью листьев.
У ряда растений развита гетерофилия (разнолистность). У кувшинок и кубышек плавающие листья сильно отличаются от погруженных – их верхняя поверхность плотная и кожистая, с большим количеством устьиц, что способствует лучшему газообмену с воздухом, на нижней стороне устьиц нет.
Из-за низкой температуры воды, отрицательно влияющей на органы размножения, и высокой плотности среды, затрудняющей перенос пыльцы, погруженные в воду растения размножаются вегетативным путем. Однако многие из них выносят цветоносные стебли в воздушную среду и размножаются половым путем. Их пыльца, плоды и семена разносятся ветром и поверхностными течениями. Поверхностные течения используют и прибрежные растения. Их плоды обладают высокой плавучестью и могут длительное время, находясь в воде, не терять всхожести.
Адаптивные особенности водных животных. Адаптации животных к водной среде более разнообразны, чем растений. Для животных, обитающих в толще воды, характерны, прежде всего, приспособления, увеличивающие их плавучесть и позволяющие им противостоять движению воды и течениям. У мелких форм наблюдается редукция скелетных образований. Они имеют пористые раковины или полые внутри иглы скелетов. Удельная плотность тела уменьшается за счет наличия воды, воздуха или жира в тканях.
Для пассивно плавающих в толще воды животных характерно также увеличение удельной поверхности тела. Это достигается уплощением тела, образованием всевозможных шипов, выростов.
Активное плавание осуществляется с помощью ресничек, жгутиков, а также изгибания тела. Получило распространение плавание реактивным способом за счет энергии выбрасываемой струи воды. Так некоторые кальмары развивают скорость 40–50 км/час. У более крупных животных имеются специализированные конечности – плавники, ласты. Тело у таких животных покрыто слизью и имеет обтекаемую форму.
Пресноводные животные при передвижении используют поверхностную пленку воды. По ней свободно бегают жуки-вертячки, клопы-водомерки. Их покровы не смачиваются водой, а конечности имеют особое строение.
Донные организмы, наоборот, вырабатывают приспособления, уменьшающие плавучесть и позволяющие удерживаться на дне даже в быстротекущих водах. Хорошо известны тяжелые раковины моллюска тридакны, которые свободно лежат на малой глубине и удерживаются на рифах благодаря своей массе.
Только в водной среде встречаются животные, ведущие неподвижный образ жизни. Из прикрепленных к грунту наиболее известны губки, гидроидные и коралловые полипы, морские лилии, двустворчатые моллюски и др.
Для водных животных имеет значение давление среды. Среди эврибатных – обитающих и при высоком и при низком давленях – выделяются голотурии, живущие на глубине от 100 до 9000 м. Среди стенобатных – морские лилии, погонофоры, обитающие на глубинах от 3000 до 10 000 м. Для глубоководных животных характерно слабое развитие или отсутствие известкового скелета, редукция органов зрения, усиление развития осязательных рецепторов, отсутствие пигментации тела или, наоборот, темная окраска.
|
Черного моря (В) с индексами атмосферной
Таблица 4
| № п/п
| Предикторы
| Сдвиг
(годы)
| Коэф.
корр.
| Вид
корр.
| Уровни
значим.
| Уравнения
регрессии
| |
| 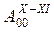
|
| 0, 58
| Парн.
| 0, 01
| В=15, 5 -4480,
| |
| 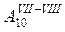
|
| 0, 58
| Парн.
| 0, 01
| В=97, 5 +324, 8
| |
| 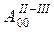
|
| 0, 73
| Парн.
| 0, 01
| В=23, 4 +255, 0
| |
| 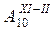
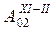
|
| 0, 69
| Множ.
| 0, 01
|
|
Мы видим, что самые высокие коэффициенты корреляции получены при сдвиге в 6 и 12 лет. В биотической и абиотической частях экосистем не обнаружены признаки «памяти» со сроком в 6 лет. Следовательно, такой уровень статистической связи может быть признаком наличия периодичности в атмосферной циркуляции, что и отражается на колебаниях в живой части экосистемы. Указанные ритмы и были выявлены с помощью периодограмманализа. Таким образом, 3-е уравнение таблицы 4 мы можем использовать для ориентировочной оценки уровня первичной продуктивности с 6-летней заблаговременностью, с учетом того, что обеспеченность прогноза при проверке на независимом материале составила 83%.
Корреляционный анализ показал зависимость урожайности двух основных промысловых видов рыб – анчоуса (хамсы) и шпрота от стока Днепра, поскольку его воды в значительной степени определяют биопродуктивность акватории северо-западного шельфа Черного моря.
Состояние донных биоценозов в указанном регионе зависит от кислородного режима придонных горизонтов, в сущности от уровня (площади, S) развития обычных здесь летних заморов. Прогноз этой площади с годовой заблаговременностью может быть дан по уравнению, где в качестве предикторов использованы значения солнечной активности (W), которые могут быть проэкстраполированы с учетом 11-летней цикличности, а также известный или предполагаемый уровень антропогенного отъема речного стока (q, км2):
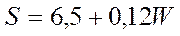
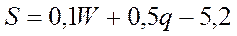
Уровень заморов и гипоксии определяет снижение биомассы филлофоры на поле Зернова, состояние и запас мидий, количество гибнущей рыбы в районах побережий на северо-западе Черного моря.
В предшествующем подразделе изложена методика прогноза состояния крилевых скопления в море Содружества с заблаговременностью в 1-3 месяца. По другим параметрам атмосферной циркуляции, в частности по аномалии повторяемости 1-го типа в марте, оказался возможным прогноз с годовой заблаговременностью следующих промыслово-биологических показателей: средневзвешенной биомассы криля на площади промысловых скоплений (В) в феврале-марте, общей его биомассы в млн. т (В1) и запаса в промысловых скоплениях (В2):
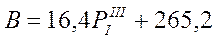
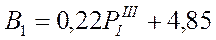
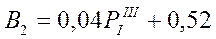
В соответствии с описанием типов атмосферной циркуляции 1-й из них отражает зональный перенос вод, что не способствует образованию промысловых скоплений. Однако, в конце полярного лета (март) он каким-то образом способствует урожайности этого вида, возможно посредством сохранения личинок криля, благодаря чему в промысловый сезон последующего года образуется повышенный запас.
В атлантическом секторе Антарктики увеличение концентраций криля в промысловых скоплениях в районах островов Мордвинова, Южных Оркнейских и острова Южная Георгия происходит при меридиональном типе циркуляции типа Мв. Его повторяемость, как было установлено, связана с уровнем солнечной активности (W). Это происходит как при экстремально высоком, так и низком значении числа Вольфа, то есть прямая связь имеет место с модулем 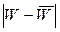 . Таким образом, ориентировочная оценка годового вылова криля в этом . Таким образом, ориентировочная оценка годового вылова криля в этом
регионе может быть сделана по году-аналогу из ряда вылова флота Украины за 1965-1995 гг в тоннах:
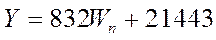
где 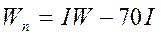 , а W – экстраполированное на текущий год число Вольфа. , а W – экстраполированное на текущий год число Вольфа.
Министерство аграрной политики украины
Керченский государственный морской технологический университет
Течения Индийского океана.
Выполнил: ст.гр. СВ-52
Першко Николай
Шифр: 12ксв1155
Керчь, 2013 г.
Вопрос 4Течения Индийского океана
|
