
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Осмос и осмотическое давление.
Осмотическое давление - это важное коллигативное свойство живых систем. Коллигативными называются свойства растворов, зависящее только от концентрации растворенного вещества, но не от его химической структуры. Осмотическое давление появляется в случае разделения двух растворов мембраной, непроницаемой для растворенных веществ. Чтобы продемонстрировать роль полупроницаемой мембраны в создании осмотического давления, представим, что 1, 0 М водный раствор сахарозы осторожно подслоен под 0, 01 М водный раствор сахарозы. В этом случае будет происходить суммарная диффузия молекул воды из раствора с низкой концентрацией сахарозы в раствор с ее высокой концентрацией и диффузия сахарозы в обратном направлении. Если эти же два раствора поместить в сообщающиеся сосуды, разделенные только жесткой мембраной, которая пропускает молекулы воды, но не сахарозы, то молекулы воды по-прежнему будут диффундировать из раствора, в котором их концентрация больше (0, 01 М раствор сахарозы), в 0, 1 М раствор, где концентрация воды меньше. Сахароза же диффундировать не сможет, так как мембрана для нее не проницаема. В результате будет происходить диффузия воды через мембрану. Такое движение воды вдоль ее концентрационного градиента называется осмосом. В результате осмоса, уровень жидкости в сосуде с большей концентрацией сахарозы, будет повышаться до тех пор, пока гидростатическое давление в этом сосуде не станет достаточным для выталкивания молекул воды в направлении против градиента концентрации с той же скоростью с какой заставляет ее двигаться осмос. Осмотическим давлением раствора называют то наименьшее давление, которое необходимо приложить к раствору для того, чтобы предотвратить перетекание к нему растворителя через полупроницаемую мембрану. Осмотическое давление, которое обозначается греческой буквой π пропорционально не только концентрации С (количество молей растворенного вещества в 1 л воды), но и абсолютной температуре Т. π = nRT/V = RTC где n - число молей растворенного вещества, R - коффициэнт пропорцианальности, называемый газовая постоянная (0, 082 л атм/моль град), V - объем в литрах. 1 молярный раствор идеального компонента (т.е. такого у которого частицы не диссоциируют и не ассоциируют) при стандартной температуре (0˚ С) покажет осмотическое давление 22, 4 атм. Поскольку коллигативные свойства зависят от суммарного количества растворенных частиц, приходящихся на единицу объема растворителя, осмотическое давление 10 мМ раствора NaCl (электролита диссоциирующего при такой концентрации на 90%) и 20 мМ раствора сахарозы почти эквивалентны. Чтобы учесть эффект диссоциации растворенных молекул осмотические свойства раствора в физиологии часто характеризуют величиной осмолярность. 1 Осмоль соответствует 6, 022х1023 растворенных в растворе частиц. Для недиссоциирующего вещества один осмоль равен одному молю, а осмолярность - молярности. Осмолярность плазмы крови и внутренней среды организма в целом составляет в норме около 300 мосмоль/л. Два раствора, в которых создается одинаковое осмотическое давление, называются изоосмотическими. Если же в одном из растворов осмотическое давление меньше то он называется гипоосмотическим, а в противном случае - гиперосмотическим по отношению к другому. Все растворы, содержащие в единице объема одинаковое количество частиц, являются изоосмотическим. Перемещение воды между двумя растворами, разделенными идеальной мембраной (т.е. пропускающей только воду) будет всегда направлено от гипоосмотического раствора к гиперосмотическому. Однако биологические мембраны не являются идеальными и в той или иной степени проницаемы для различных ионов. Если мембрана разделяет два изоосмотичных раствора различных веществ, например, NaCl и KCl и при этом пропускает, например только ионы К+, эти ионы перемещаясь по концентрационному градиенту в раствор содержащий первоначально NaCl сделают его гиперосмотичным, что в свою очередь приведет к перемещению воды от раствора KCl к раствору NaCl. Сказанное можно проиллюстрировать следующим примером. Эритроциты крови не меняются в объеме, будучи помещенные в раствор NaCl, изоосмотичный плазме крови, но набухает в растворе мочевины, также изоосмотичном плазме. Это различие обусловлено значительно более высокой способностью мочевины проникать и накапливаться во внутренней среде эритроцитов по сравнению с NaCl. В результате, по мере нагружения клетки мочевиной в нее под действием осмотического давления проникает все больше молекул воды, она набухает и может даже разрушиться. Для характеристики осмотического влияния конкретных растворов на конкретные живые клетки или ткани, используется понятие тоничность. Тоничность не является количественным параметром, а определяется экспериментально по реакции клеток или тканей на погружение их в исследуемый раствор. В отличие от осмотического давления, тоничность раствора зависит не только от концентрации растворенного вещества, но и от скорости его проникновения внутрь клеток. Осмотичность раствора и его тоничность совпадают только в том случае если растворенное вещество плохо проникает в клетки или ткани (Vпроникновения = 0). Раствор называется изотоническим по отношению к данной клетке или ткани, если клетка или ткань, погруженные в него не набухают и не сжимаются. Если ткань набухает, раствор называют гипотоничным по отношению к ткани, а если сжимается, то гипертоничным. Таким образом, об изотоничности, гипотоничности и гипертоничности имеет смысл говорить лишь по отношению конкретной системе: раствор-живые клетки (или ткань). Свойство изотоничности особенно важно учитывать при приготовлении растворов для внутривенных инъекций, в противном случае может произойти набухание и лизис эритроцитов (гемолиз). Обычно при внутривенных инъекциях лекарственный препарат вводят в изотоническом для клеток крови 0, 9% растворе NaCl на дистиллированной воде, получившем название физиологический раствор. Поддержание постоянства осмотического давления является одним из важнейших элементов гомеостаза, как организма в целом, так и составляющих его систем. На клеточном уровне осмотический гомеостаз поддерживается благодаря активному транспорту и функционированию мембранных насосов, откачивающих из клетки ионы натрия и обеспечивающих поддержание осмотического равновесия между цитозолем и внеклеточной жидкостью. В норме содержимое клеток незначительно гиперосмотично по отношению к внеклеточной среде, что обеспечивает небольшое внутриклеточное давление или тургор, поддерживающий форму клеток. Нарушение (ингибирование) активного транспорта ведет к возрастанию внутриклеточной концентрации натрия, внутриклеточное содержимое становится сильно гиперосмотично по отношению к внешней среде. В клетку начинает интенсивно поступать вода, что ведет к набуханию, деформации клетки и даже разрыву клеточной мембраны. В случае эритроцитов такой процесс получил название осмотический гемолиз. На уровне организма важную роль в поддержании осмотического или водно-электролитного гомеостаза играют специальные осморецепторы, способные определять существующее в организме и его составляющих осмотическое давление и почки, через которые выделяются из организма, как избыток воды, так и постоянно накапливающиеся осмоактивные вещества. В среднем за сутки почками выводится около 1200 мосмоль различных соединений и примерно 1 л воды. Еще около 900 мл воды в течение суток испаряется с поверхности кожи и выделяется через легкие с выдыхаемым воздухом. Основными источниками поступление воды в организм является питьевая вода и напитки (примерно 800 мл), вода в составе твердой пищи (примерно 900 мл) и вода образующаяся как продукт окисления питательных веществ (300 мл). Относительное содержание воды в организме в норме колеблется в пределах ± 0, 22 %. Так как тело человека примерно на 70 % состоит из воды, то такое колебание составляет примерно 150 мл. Потеря жидкости, превышающая 150 мл, вызывает чувство жажды и стремление пить воду, причем адоптации к этому чувству не происходит. Различают несколько видов жажды. Гиперосмотическая жажда вызывается абсолютной потерей воды, например при сильном потоотделении, или ее относительным дефицитом после употребления соленой пищи. В этом случае возрастает осмомолярность внеклеточной жидкости, что приводит к выходу воды и осмотическому сжатию клеток центральных осморецепторов, расположенных в отделе ЦНС и сигнализирующих о недостатке воды в организме. При уменьшении общего объема жидкости, без изменения ее осмотической концентрации, например при донорской сдаче крови, развивается гиповолемическая жажда. В этом случае о нарушении водно-солевого баланса в организме сигнализирует другой тип рецепторов. Мембранный потенциал. Разность электрических потенциалов (в вольтах или мв) между жидкостью, находящейся по одну сторону мембраны и жидкостью по другую ее сторону называется мембранным потенциалом (МП) и обозначается Vм. Величина МП живых клеток составляет обычно от -30 до -100 мв и вся эта разность потенциалов создается в областях непосредственно прилегающих с обоих сторон к клеточной мембране. Уменьшение величины МП называют деполяризацией, увеличение - гиперполяризацией, восстановление исходного значения после деполяризации - реполяризация. Мембранный потенциал существует во всех клетках, но в возбудимых тканях (нервных, мышечных, железистых), мембранный потенциал или как его еще называют в этих тканях, мембранный потенциал покоя, играет ключевую роль в реализации их физиологических функций. Мембранный потенциал обусловлен двумя основными свойствами всех эукариотических клеток: 1) асимметричным распределением ионов между вне- и внутриклеточной жидкостью, поддерживаемым метаболическими процессами; 2) Избирательной проницаемостью ионных каналов клеточных мембран. Чтобы уяснить себе как возникает МП представим себе, что некий сосуд разделен на два отсека мембраной, проницаемой только для ионов калия. Пусть в первом отсеке содержится 0, 1 М, а во втором 0, 01 М раствор КСl. Поскольку концентрация ионов калия (К+) в первом отсеке в 10 раз выше, чем во втором, то в начальный момент на каждые 10 ионов К+ диффундирующих из отсека 1 во второй будет приходится один ион диффундирующий в обратном направлении. Так как анионы хлора (Сl-) не могут переходить через мембрану вместе с катионами калия, то во втором отсеке будет образовываться избыток положительно заряженных ионов и, напротив, в отсеке 1 окажется избыток ионов Сl-. В результате возникает трансмембранная разность потенциалов, препятствующая дальнейшей диффузии К+ во второй отсек, поскольку для этого им нужно преодолеть притяжение отрицательных ионов Сl-, в момент вхождения в мембрану со стороны отсека 1 и отталкивание одноименных ионов на выходе из мембраны в отсек 2. Таким образом, на каждый ион К+, проходящий через мембрану в этот момент действуют две силы - химический градиент концентраций (или химическая разность потенциалов), способствующая переходу ионов калия из первого отсека во второй, и электрическая разность потенциалов, заставляющая ионы К+ двигаться в обратном направлении. После того как эти две силы уравновесятся, количество ионов К+ перемещающееся из отсека 1 в отсек 2 и обратно сравняется, установится электрохимическое равновесие. Соответствующая такому состоянию трансмембранная разность потенциалов называется равновесным потенциалом, в данном конкретном случае равновесным потенциалом для ионов калия (Ек). В конце 19 века Вальтер Нернст установил, что равновесный потенциал зависит от абсолютной температуры, валентности диффундирующего иона и от отношения концентраций данного иона по разные стороны мембраны: Ex = где Ех- равновесный потенциал для иона X, R - универсальная газовая постоянная = 1, 987 кал/(моль град), T- абсолютная температура в градусах Кельвина, F - число Фарадея = 23060 кал/в, Z - заряд переносимого иона, [X]1 и [X]2 - концентрации иона в отсеках 1 и 2. Если перейти от натурального логарифма к десятичному, то для температуры 18˚ С и моновалентного иона можно записать уравнение Нернста следующим образом: Ех= 0, 058 Рассчитаем с помощью уравнения Нернста калиевый равновесный потенциал для воображаемой клетки, приняв, что внеклеточная концентрация калия [К+]н= 0, 01 М, а внутриклеточная - [К+]в = 0, 1 М: Ек= 0, 058
В данном случае, Екотрицателен, поскольку ионы калия будут выходить из гипотетичной клетки, заряжая отрицательно слой цитоплазмы, прилегающий к внутренней стороне мембраны. Поскольку в данной гипотетичной системе имеется только один диффундирующий ион, то калиевый равновесный потенциал будет равен мембранному потенциалу (Ек= Vм).
Приведенный механизм ответственен и за образование мембранного потенциала в реальных клетках, но в отличие от рассмотренной упрощенной системы, в которой через " идеальную" мембрану мог диффундировать только один ион, реальные клеточные мембраны пропускают в той или иной все неорганические ионы. Однако, чем менее мембрана проницаема для какого-либо иона, тем меньшее влияние он оказывает на МП. Учитывая это обстоятельство, Голдманом в 1943г. было предложено уравнение для расчета величины МП реальных клеток, учитывающее концентрации и относительную проницаемость через плазматическую мембрану всех диффундирующих ионов: Vм = 0, 058 Используя метод меченых изотопов, Ричард Кейнс в 1954 г. определил проницаемость клеток мышц лягушки для основных ионов. Оказалось, что проницаемость для натрия примерно в 100 раз меньше, чем для калия, а ион Сl-не вносит заметного вклада в создание МП мышечных клеток. Поэтому для мембран мышечных клеток уравнение Голдмана можно записать в следующем упрощенном виде: Vм = 0, 058 Vм = 0, 058
Исследования с применением вводимых в клетки микроэлектродов, показали, что потенциал покоя клеток скелетных мышц лягушки колеблется от -90 до -100 мв. Такое хорошее соответствие экспериментальных данных теоретическим подтверждает, что потенциал покоя определяется диффузионными потоками неорганических ионов. При этом, в реальных клетках мембранный потенциал близок к равновесному потенциалу иона, который характеризуется максимальной трансмембранной проницаемостью, а именно к равновесному потенциалу иона калия. Роль кальция Лекция 3 Основы биоэнергетики. Химическая энергия, особенности ее образования и утилизации в живых системах. Молекулярные механизмы преобразования энергии в живых клетках изучает биоэнергетика. Восстановленные органические соединения, служащие источником энергии живым организмам, окисляются до СО2 и Н2О, а высвобождающаяся энергия используется для синтеза АТФ и выполнения различных работ. Прежде чем перейти к рассмотрению конкретных биоэнергетических процессов в клетке вспомним, что в соответствии со вторым законом термодинамики энтропия любой замкнутой системы с течением времени может только возрастать. Энтропия - термодинамическая категория, характеризующая степень неупорядоченности системы. Поэтому, возрастание энтропии, по сути, есть увеличение степени неупорядоченности системы, т.е. переход от более упорядоченной структуры к менее упорядоченной и снижение доли полезной энергии (т.е. энергии, с помощью которой может быть совершена работа). Другими словами из второго закона термодинамики следует, что в любой изолированной системе степень неупорядоченности может только возрастать, а величина полезной (свободной) энергии - уменьшаться. В тоже время живые организмы, по мере того как проходят развитие от оплодотворенной яйцеклетки до состояния взрослой особи, постоянно усложняют свою структуру. Казалось бы, в живых системах второй закон термодинамики не выполняется. Однако, следует иметь ввиду, что этот закон относится к замкнутой системе, а живой организм является открытой системой, для которых характерен постоянный обмен с окружающей средой веществом и энергией и поддерживание относительно низкого уровня энтропии, за счет повышения ее в окружающей среде. Способность извлекать полезную энергию из окружающей среды является одним из тех замечательных свойств, которые отличают живые организмы от неживой природы. Потребляя растительную или животную пищу, живой организм превращает содержащиеся в ней высокоорганизованные молекулы в СО2, Н2О и низкомолекулярные азотсодержащие соединения, имеющие значительно более высокий уровень энтропии, а образующуюся при этом энергию использует, в том числе на развитие и рост собственного организма, сопровождающийся понижением энтропии. Первичным же источником энергии на земле для большинства организмов является солнце, энтропия которого постоянно повышается и только незначительная часть выделяемой при этом энергии улавливается биосферой и утилизируется растениями и некоторыми бактериями в реакциях фотосинтеза, а остальная безвозвратно теряется во Вселенной. Даже открытая недавно обильная жизнь глубоко на дне океанов в области океанических трещин на т.н. «черных курильщиках» зависит от солнца, поскольку является аэробной, а кислород берется в атмосфере и океане только благодаря процессу фотосинтеза. В процессе эволюции в организме животных и естественно человека сформировалось множество путей получения, транспорта и утилизации энергии, и в подавляющем большинстве из них для аккумуляции химической энергии используются специальные соединения – фосфагены, имеющие в своей структуре так называемую высокоэнергетическую или макроэргическую связь. Аденозин-5'-трифосфат (АТФ) представляет собой наиболее распространенный и важнейший представитель этой группы соединений. Кроме АТФ, высокоэнергетические связи имеют ряд других органических полифосфатов, например ГТФ, ЦТФ Для обозначения макроэргической связи используется символ ~. Так молекулу АТФ сокращенно обозначают А-Р-Ф~Ф~Ф (А-аденин, Р-рибоза). Некоторые бактерии даже запасают энергию в форме неорганического полиметафосфата – линейного полимера неопределенной длинны. При рН=7, 0 АТФ представляет анион с высоким зарядом. Трифосфатная группа содержит 4 ОН группы способные к ионизации. 3 из них имеют низкое значение рК и при рН=7 полностью диссоциируют, четвертая группа диссоциирует на 75%. Высокая концентрация отрицательных зарядов является важным фактором, обуславливающим высокоэнергетическую природу АТФ. Эти заряды располагаются близко друг к другу и между ними существует сильное отталкивание. При гидролитическом отщеплении концевой фосфатной группы силы отталкивания ослабевают. Образовавшиеся продукты не стремятся воссоединиться. Вторая важная особенность, обуславливающая большую отрицательную величину Δ G' гидролиза АТФ заключается в том, что оба продукта гиролиза стабилизированы за счет сопряжения. Впрочем, в клетках АТФ почти не встречается в форме свободных ионов, а чаще в виде комплекса с ионами Mg или Mn. Понятие энергии фосфатной связи, которым пользуются биохимики не следует путать с понятием энергии связи, под которой в физической химии понимают энергию необходимую для разрыва в молекуле связи между 2 атомами. Под энергией макроэргической фосфатной связи понимают разность между свободной энергией исходных веществ и свободной энергией продуктов реакции, в процессе которой фосфорилированное соединение гидролизуется с образованием неорганического фосфата. Перенос концевой фосфатной группы АТФ на воду сопровождается освобождением свободной энергии, примерно вдвое большей, чем при гидролизе обычной фосфоэфирной связи, например в аденозинмонофосфате. Свободная энергия характеризует ту долю полной энергии заключенной в веществе, с помощью которой может быть совершена работа в изотермических и изобарических условиях, т.е. при постоянной температуре и давлении, а именно такие условия характерны для биологических систем. Изменение свободной энергии измеряется в ккал/моль и обозначается Δ G' если процесс протекает в стандартных условиях (рН=7, Т=25 º С, субстраты и продукты реакции взяты в одинаковой концентрации равной 1 моль/л) и символом Δ G для реакции в конкретных условиях реальных биологических систем. В случае гидролиза АТФ, Δ G' = -7 ккал/моль, а Δ G = -11-13 ккал/моль Живые организмы используют энергию АТФ для выполнения механической работы мышц, создания концентрационных градиентов и трансмембранных электрических потенциалов. Конкретные механизмы, обеспечивающие протекание данных процессов будут изложены в последующих лекциях, а теперь рассмотрим, общие принципы, обеспечивающие возможность использования энергии АТФ для совершения химической работы. Представим себе типичный биосинтетический, т.е. анаболический процесс (анаболизм от греч. подъем те процессы синтеза, построения структурных частей клетки, противоположен катаболизму), при котором два мономера -Х и Y- должны соединиться друг с другом в реакции дегидратации (называемой также конденсацией), сопровождающейся выделением воды: Х-ОН + Y-Н Большинство таких реакций являются эндергоническими (не путать с эндотермическими) и сопровождаются возрастанием свободной энергии на величинуΔ G, а движущей силой любой химической реакции является уменьшение свободной энергии и самопроизвольно протекают только экзергонические реакции и, следовательно, почти всегда предпочтительнее обратная катаболическая реакция (называемая гидролизом), поскольку в этом случае имеет место выделение свободной энергии равное по абсолютной величине ее потреблению в реакции конденсации. Поэтому в клетке такие реакции протекают в несколько стадий, в результате которых происходит сопряжение энергетически невыгодного синтеза с сильно экзергоническиой реакцией, протекающей с участием АТФ. Механизм сопряжения реализуется следующим образом. Реакции синтеза, приведенной выше и сопровождающейся возрастанием свободной энергии (предположим Δ G' = 3 ккал/моль), предшествует фосфорилирование одного из субстратов молекулой АТФ. А-Р-Ф~Ф~Ф + ХОН → А-Р-Ф~Ф + Х-О-Ф - Δ G = 4 ккал/моль Δ G' этой реакции значительно меньше (4 ккал/моль), чем реакции гидролиза АТФ и переноса фосфатной группы на воду (примерно 11 ккал/моль). Остальная энергия (7 ккал/моль) аккумулируется в молекуле активированного субстрата, который вовлекается в последующую синтетическую реакцию. Однако, теперь свободная энергия исходных веществ выше, чем свободная энергия продуктов реакции (7 - 3 = 4 ккал/моль) и, реакция протекает слева направо, т.е. в направлении синтеза, сопровождаясь уменьшением свободной энергии: Х-О-Ф + Y-Н → X-Y + Фн - Δ G = ( 4 ккал/моль) Таким образом, за счет накопленной в молекуле АТФ энергии эндергоническая реакция замещается двумя сопряженными экзергоническими реакциями. Другой путь использования АТФ для совершения химической работы реализуется через последовательно сопряженные или как их называют тандемные реакции. В этом случае эндергоническая реакция протекает благодаря следующей за ней экзергонической реакции. Возможность протекания тандемных реакций обусловлена тем обстоятельством, что увеличение свободной энергии не исключает протекание химической реакции, а лишь указывает на то, что величина константы равновесия такой реакции меньше единицы, т.е. если имеет место реакция:
А + B
то Кeq = и, следовательно, равновесие реакции смещено влево и это смещение тем более выражено, чем больше по абсолютной величине Δ G'. Однако если за реакцией 1 следует реакция 2 С +X и для этой реакции величина Δ G' сильно отрицательна, то промежуточный продукт реакции С будет потребляться, смещая при этом равновесие реакции 1 вправо. В том случае если суммарная свободная энергия двух реакций будет величиной отрицательной они будут протекать в направлении потребления субстратов А и В и образования продукта СХ. Теперь рассмотрим где в клетке локализованы механизмы получения энергии и перевода фосфагенов из " незаряженной" формы (например АДФ) в " заряженную" (АТФ) и как они функционируют. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 657; Нарушение авторского права страницы

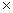 lg
lg 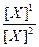
 = 0, 058
= 0, 058  =0, 058
=0, 058 


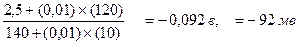

 X-Y + Н2О + Δ G (3 ккал/моль)
X-Y + Н2О + Δ G (3 ккал/моль)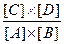 < 1
< 1